3.8.1. The female system
The main functions of the female reproductive system are egg production, including the provision of a protective coating in many insects, and the storage of the male’s spermatozoa until the eggs are ready to be fertilized. Transport of the spermatozoa to the female’s storage organ and their subsequent controlled release requires movement of the spermatozoa, which in some species is known to be mediated by muscular contractions of parts of the female reproductive tract.
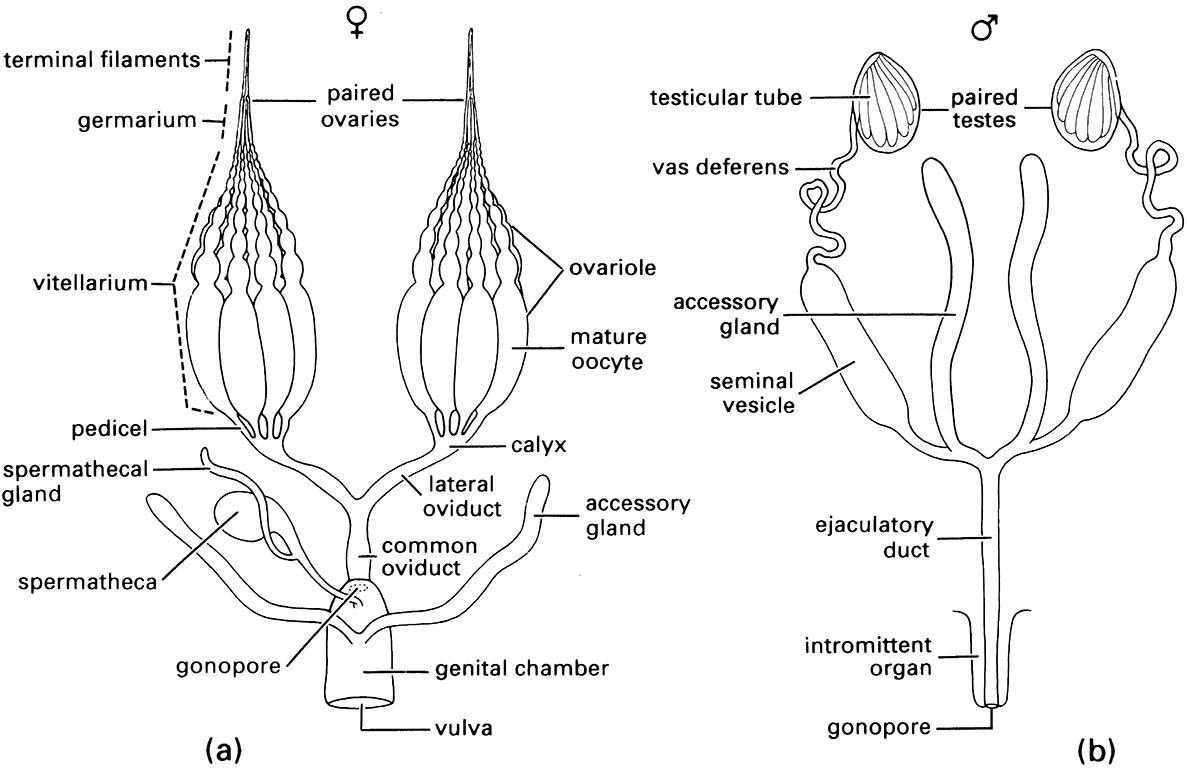
The basic components of the female system (Fig. 3.20a) are paired ovaries, which empty their mature oocytes (eggs) via the calyces (singular: calyx) into the lateral oviducts, which unite to form the common (or median) oviduct. The gonopore (opening) of the common oviduct usually is concealed in an inflection of the body wall that typically forms a cavity, the genital chamber. This chamber serves as a copulatory pouch during mating and thus often is known as the bursa copulatrix. Its external opening is the vulva. In many insects the vulva is narrow and the genital chamber becomes an enclosed pouch or tube, referred to as the vagina. Two sorts of ectodermal glands open into the genital chamber. The first is the spermatheca, which stores spermatozoa until needed for egg fertilization. Typically, the spermatheca is single, generally sac-like with a slender duct and often has a diverticulum that forms a tubular spermathecal gland. The gland or glandular cells within the storage part of the spermatheca provide nourishment to the contained spermatozoa. The second type of ectodermal gland, known collectively as accessory glands, opens more posteriorly in the genital chamber and has a variety of functions depending on the species (see section 5.8).
Each ovary is composed of a cluster of ovarian or egg tubes, the ovarioles, each consisting of a terminal filament, a germarium (in which mitosis gives rise to primary oocytes), a vitellarium (in which oocytes grow by deposition of yolk in a process known as vitellogenesis; section 5.11.1), and a pedicel (or stalk). An ovariole contains a series of developing oocytes, each surrounded by a layer of follicle cells forming an epithelium (the oocyte and its epithelium is termed a follicle); the youngest oocytes occur near the apical germarium and the most mature near the pedicel. Three different types of ovariole are recognized based on the manner in which the oocytes are nourished. A panoistic ovariole lacks specialized nutritive cells so that it contains only a string of follicles, with the oocytes obtaining nutrients from the hemolymph via the follicular epithelium. Ovarioles of the other two types contain trophocytes (nurse cells) that contribute to the nutrition of the developing oocytes. In a telotrophic (or acrotrophic) ovariole the trophocytes are confined to the germarium and remain connected to the oocytes by cytoplasmic strands as the oocytes move down the ovariole. In a polytrophic ovariole a number of trophocytes are connected to each oocyte and move down the ovariole with it, providing nutrients until depleted; thus individual oocytes alternate with groups of successively smaller trophocytes. Different suborders or orders of insects usually have only one of these three ovariole types.
Accessory glands of the female reproductive tract often are referred to as colleterial or cement glands because in most insect orders their secretions surround and protect the eggs or cement them to the substrate (section 5.8). In other insects the accessory glands may function as poison glands (as in many Hymenoptera) or as “milk” glands in the few insects (e.g. tsetse flies, Glossina spp.) that exhibit adenotrophic viviparity (section 5.9). Accessory glands of a variety of forms and functions appear to have been derived independently in different orders and even may be non-homologous within an order, as in Coleoptera.
(After Snodgrass 1935)

