3.1.2. Muscle attachments
Vertebrates’ muscles work against an internal skeleton, but the muscles of insects must attach to the inner surface of an external skeleton. As musculature is mesodermal and the exoskeleton is of ectodermal origin, fusion must take place. This occurs by the growth of tonofibrillae, fine connecting fibrils that link the epidermal end of the muscle to the epidermal layer (Fig. 3.2a,b). At each molt tonofibrillae are discarded along with the cuticle and therefore must be regrown.
At the site of tonofibrillar attachment, the inner cuticle often is strengthened through ridges or apodemes, which, when elongated into arms, are termed apophyses (Fig. 3.2c). These muscle attachment sites, particularly the long, slender apodemes for individual muscle attachments, often include resilin to give an elasticity that resembles that of vertebrate tendons.
Some insects, including soft-bodied larvae, have mainly thin, flexible cuticle without the rigidity to anchor muscles unless given additional strength. The body contents form a hydrostatic skeleton, with turgidity maintained by criss-crossed body wall “turgor” muscles that continuously contract against the incompressible fluid of the hemocoel, giving a strengthened foundation for other muscles. If the larval body wall is perforated, the fluid leaks, the hemocoel becomes compressible and the turgor muscles cause the larva to become flaccid.
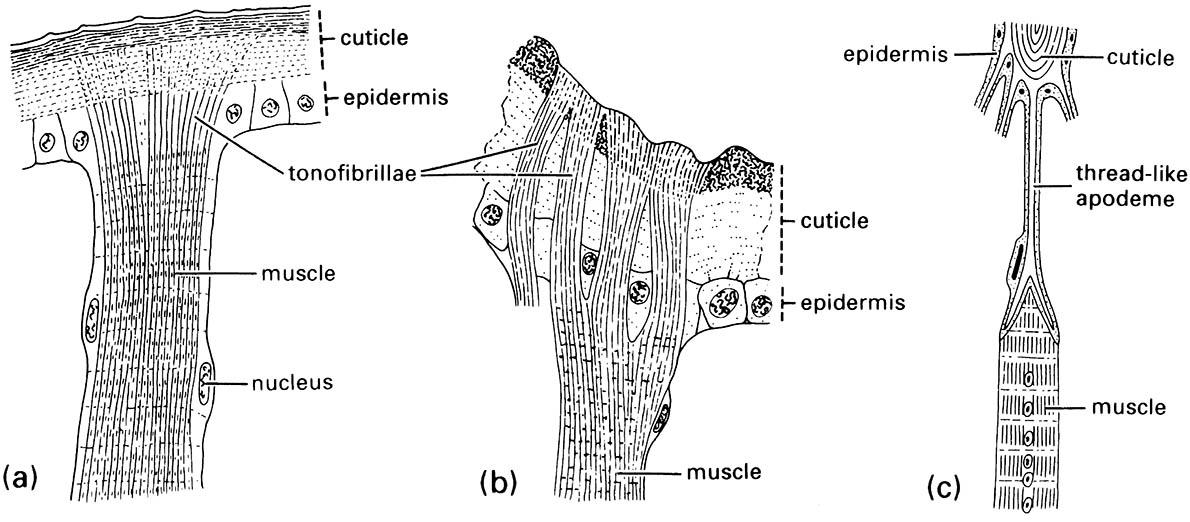
(a) tonofibrillae traversing the epidermis from the muscle to the cuticle; (b) a muscle attachment in an adult beetle of Chrysobothrus femorata (Coleoptera: Buprestidae); (c) a multicellular apodeme with a muscle attached to one of its thread-like, cuticular “tendons” or apophyses. (After Snodgrass 1935)

