3.1.3. Crawling, wriggling, swimming, and walking
Soft-bodied larvae with hydrostatic skeletons move by crawling. Muscular contraction in one part of the body gives equivalent extension in a relaxed part elsewhere on the body. In apodous (legless) larvae, such as dipteran “maggots”, waves of contractions and relaxation run from head to tail. Bands of adhesive hooks or tubercles successively grip and detach from the substrate to provide a forward motion, aided in some maggots by use of their mouth hooks to grip the substrate. In water, lateral waves of contraction against the hydrostatic skeleton can give a sinuous, snake-like, swimming motion, with anterior-to-posterior waves giving an undulating motion.
Larvae with thoracic legs and abdominal prolegs, like caterpillars, develop posterior-to-anterior waves of turgor muscle contraction, with as many as three waves visible simultaneously. Locomotor muscles operate in cycles of successive detachment of the thoracic legs, reaching forwards and grasping the substrate. These cycles occur in concert with inflation, deflation, and forward movement of the posterior prolegs.
Insects with hard exoskeletons can contract and relax pairs of agonistic and antagonistic muscles that attach to the cuticle. Compared to crustaceans and myriapods, insects have fewer (six) legs that are located more ventrally and brought close together on the thorax, allowing concentration of locomotor muscles (both flying and walking) into the thorax, and providing more control and greater efficiency. Motion with six legs at low to moderate speed allows continuous contact with the ground by a tripod of fore and hind legs on one side and mid leg of the opposite side thrusting rearwards (retraction), whilst each opposite leg is moved forwards (protraction) (Fig. 3.3). The center of gravity of the slow-moving insect always lies within this tripod, giving great stability. Motion is imparted through thoracic muscles acting on the leg bases, with transmission via internal leg muscles through the leg to extend or flex the leg. Anchorage to the substrate, needed to provide a lever to propel the body, is through pointed claws and adhesive pads (the arolium or, in flies and some beetles, pulvilli). Claws such as those illustrated in the vignette to Chapter 2 can obtain purchase on the slightest roughness in a surface, and the pads of some insects can adhere to perfectly smooth surfaces through the application of lubricants to the tips of numerous fine hairs and the action of close- range molecular forces between the hairs and the substrate.
When faster motion is required there are several alternatives — increasing the frequency of the leg movement by shortening the retraction period; increasing the stride length; altering the triangulation basis of support to adopt quadrupedy (use of four legs); or even hind-leg bipedality with the other legs held above the substrate. At high speeds even those insects that maintain triangulation are very unstable and may have no legs in contact with the substrate at intervals. This instability at speed seems to cause no difficulty for cockroaches, which when filmed with high-speed video cameras have been shown to maintain speeds of up to 1 m s-1 whilst twisting and turning up to 25 times per second. This motion was maintained by sensory information received from one antenna whose tip maintained contact with an experimentally provided wall, even when it had a zig-zagging surface.
Many insects jump, some prodigiously, usually using modified hind legs. In orthopterans, flea beetles (Alticinae), and a range of weevils, an enlarged hind (meta-) femur contains large muscles whose slow contraction produces energy stored by either distortion of the femorotibial joint or in some spring-like sclerotization, for example the meta-tibial extension tendon. In fleas, the energy is produced by the trochanter levator muscle raising the femur and is stored by compression of an elastic resilin pad in the coxa. In all these jumpers, release of tension is sudden, resulting in propulsion of the insect into the air — usually in an uncontrolled manner, but fleas can attain their hosts with some control over the leap. It has been suggested that the main benefit for flighted jumpers is to get into the air and allow the wings to be opened without damage from the surrounding substrate.
In swimming, contact with the water is maintained during protraction, so it is necessary for the insect to impart more thrust to the rowing motion than to the recovery stroke to progress. This is achieved by expanding the effective leg area during retraction by extending fringes of hairs and spines (Fig. 10.8). These collapse onto the folded leg during the recovery stroke. We have seen already how some insect larvae swim using contractions against their hydrostatic skeleton. Others, including many nymphs and the larvae of caddisflies, can walk underwater and, particularly in running waters, do not swim routinely.
The surface film of water can support some specialist insects, most of which have hydrofuge (water-repelling) cuticles or hair fringes and some, such as gerrid water- striders (Fig. 5.7), move by rowing with hair-fringed legs.
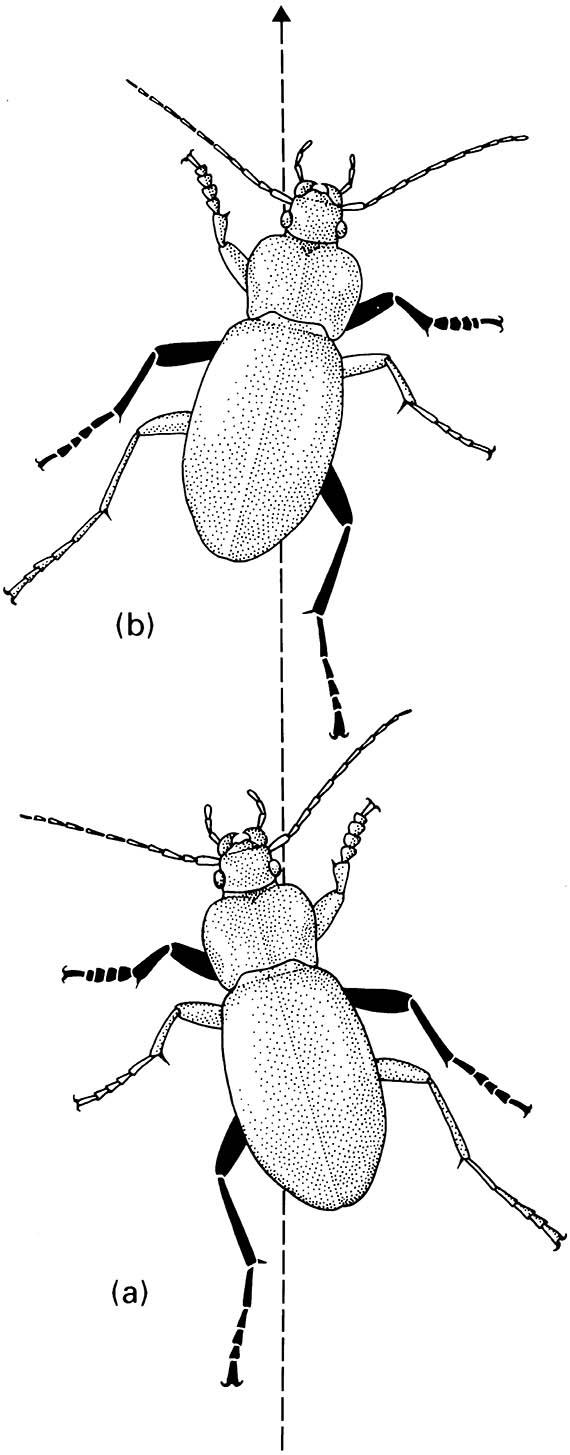
The three blackened legs are those in contact with the ground in the two positions illustrated — (a) is followed by (b). (After Wigglesworth 1972)
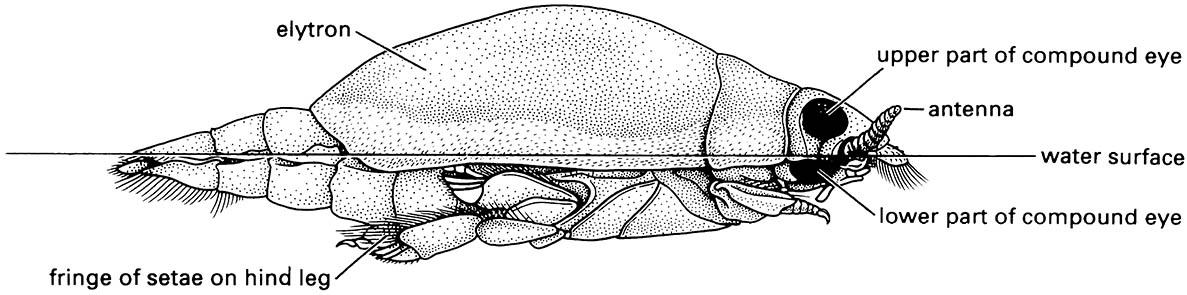
Note: the divided compound eye allows the beetle to see both above and below water simultaneously; hydrofuge hairs on the margin of the elytra repel water. (After White et al. 1984)
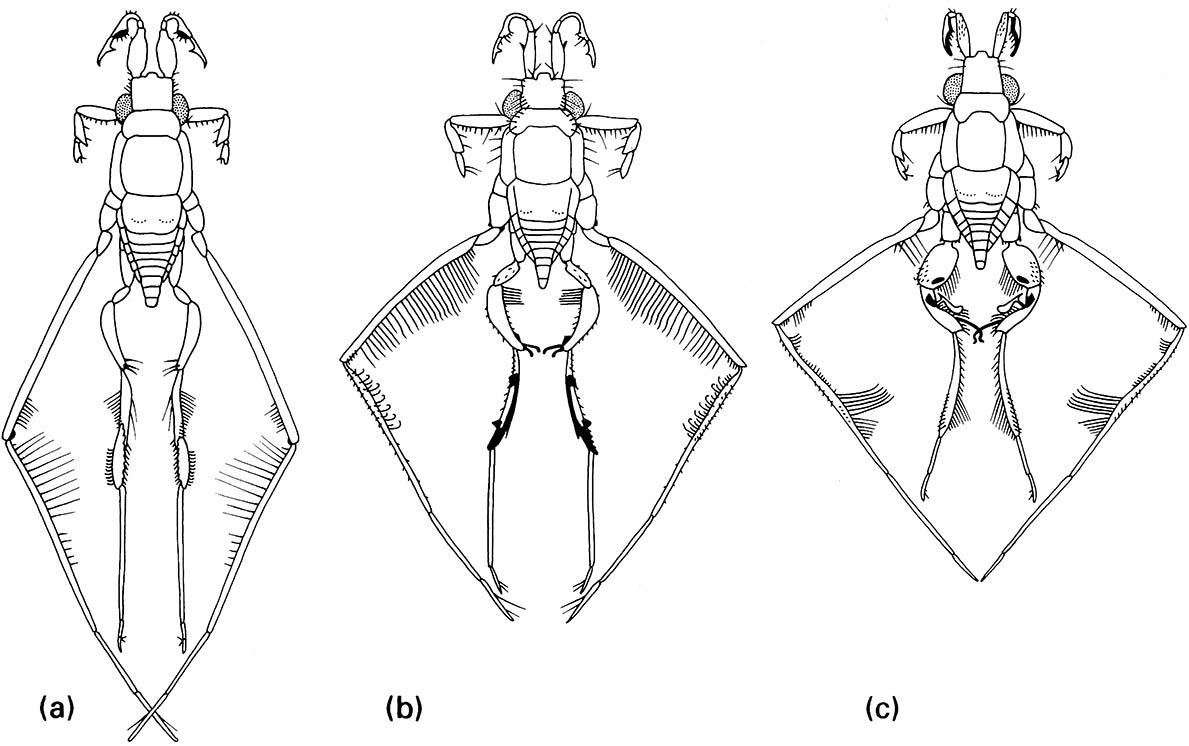
These non-genitalic male structures are specialized for contact with the female during mating, when the male rides on her back. Females of all species have a similar body form. (a) R. trulliger; (b) R. rileyi; (c) R. bergrothi. (After Hungerford 1954)

