3.1.4. Flight
The development of flight allowed insects much greater mobility, which helped in food and mate location and gave much improved powers of dispersal. Importantly, flight opened up many new environments for exploitation. Plant microhabitats such as flowers and foliage are more accessible to winged insects than to those without flight.
Fully developed, functional, flying wings occur only in adult insects, although in nymphs the developing wings are visible as wing buds in all but the earliest instars. Usually two pairs of functional wings arise dorsolaterally, as fore wings on the second and hind wings on the third thoracic segment. Some of the many derived variations are described in section 2.4.2.
To fly, the forces of weight (gravity) and drag (air resistance to movement) must be overcome. In gliding flight, in which the wings are held rigidly outstretched, these forces are overcome through the use of passive air movements — known as the relative wind. The insect attains lift by adjusting the angle of the leading edge of the wing when orientated into the wind. As this angle (the attack angle) increases, so lift increases until stalling occurs, i.e. when lift is catastrophically lost. In contrast to aircraft, nearly all of which stall at around 20°, the attack angle of insects can be raised to more than 30°, even as high as 50°, giving great maneuverability. Aerodynamic effects such as enhanced lift and reduced drag can come from wing scales and hairs, which affect the boundary layer across the wing surface.
Most insects glide a little, and dragonflies (Odonata) and some grasshoppers (Orthoptera), notably locusts, glide extensively. However, most winged insects fly by beating their wings. Examination of wing beat is difficult because the frequency of even a large slow- flying butterfly may be five times a second (5 Hz), a bee may beat its wings at 180 Hz, and some midges emit an audible buzz with their wing-beat frequency of greater than 1000 Hz. However, through the use of slowed- down, high-speed cine film, the insect wing beat can be slowed from faster than the eye can see until a single beat can be analyzed. This reveals that a single beat comprises three interlinked movements. First is a cycle of downward, forward motion followed by an upward and backward motion. Second, during the cycle each wing is rotated around its base. The third component occurs as various parts of the wing flex in response to local variations in air pressure. Unlike gliding, in which the relative wind derives from passive air movement, in true flight the relative wind is produced by the moving wings. The flying insect makes constant adjustments, so that during a wing beat, the air ahead of the insect is thrown backwards and downwards, impelling the insect upwards (lift) and forwards (thrust). In climbing, the emergent air is directed more downwards, reducing thrust but increasing lift. In turning, the wing on the inside of the turn is reduced in power by decrease in the amplitude of the beat.
Despite the elegance and intricacy of detail of insect flight, the mechanisms responsible for beating the wings are not excessively complicated. The thorax of the wing-bearing segments can be envisaged as a box with the sides (pleura) and base (sternum) rigidly fused, and the wings connected where the rigid tergum is attached to the pleura by flexible membranes. This membranous attachment and the wing hinge are composed of resilin (section 2.1), which gives crucial elasticity to the thoracic box. Flying insects have one of two kinds of arrangements of muscles powering their flight:
- direct flight muscles connected to the wings;
- an indirect system in which there is no muscle-to-wing connection, but rather muscle action deforms the thoracic box to move the wing.
A few old groups such as Odonata and Blattodea appear to use direct flight muscles to varying degrees, although at least some recovery muscles may be indirect. More advanced insects use indirect muscles for flight, with direct muscles providing wing orientation rather than power production.
Direct flight muscles produce the upward stroke by contraction of muscles attached to the wing base inside the pivotal point (Fig. 3.4a). The downward wing stroke is produced through contraction of muscles that extend from the sternum to the wing base outside the pivot point (Fig. 3.4b). In contrast, indirect flight muscles are attached to the tergum and sternum. Contraction causes the tergum, and with it the very base of the wing, to be pulled down. This movement levers the outer, main part of the wing in an upward stroke (Fig. 3.4c). The down beat is powered by contraction of the second set of muscles, which run from front to back of the thorax, thereby deforming the box and lifting the tergum (Fig. 3.4d). At each stage in the cycle, when the flight muscles relax, energy is conserved because the elasticity of the thorax restores its shape.
Primitively, the four wings may be controlled independently with small variation in timing and rate allowing alteration in direction of flight. However, excessive variation impedes controlled flight and the beat of all wings is usually harmonized, as in butterflies, bugs, and bees, for example, by locking the fore and hind wings together, and also by neural control. For insects with slower wing-beat frequencies (<100 Hz), such as dragonflies, one nerve impulse for each beat can be maintained by synchronous muscles. However, in faster-beating wings, which may attain a frequency of 100 to over 1000 Hz, one impulse per beat is impossible and asynchronous muscles are required. In these insects, the wing is constructed such that only two wing positions are stable — fully up and fully down. As the wing moves from one extreme to the alternate one, it passes through an intermediate unstable position. As it passes this unstable (“click”) point, thoracic elasticity snaps the wing through to the alternate stable position. Insects with this asynchronous mechanism have peculiar fibrillar flight muscles with the property that, on sudden release of muscle tension, as at the click point, the next muscle contraction is induced. Thus muscles can oscillate, contracting at a much higher frequency than the nerve impulses, which need be only periodic to maintain the insect in flight. Harmonization of the wing beat on each side is maintained through the rigidity of the thorax — as the tergum is depressed or relaxed, what happens to one wing must happen identically to the other. However, insects with indirect flight muscles retain direct muscles that are used in making fine adjustments in wing orientation during flight.
Direction and any deviations from course, perhaps caused by air movements, are sensed by insects predominantly through their eyes and antennae. However, the true flies (Diptera) have extremely sophisticated sensory equipment, with their hind wings modified as balancing organs. These halteres, which each comprise a base, stem, and apical knob (Fig. 2.22f ), beat in time but out of phase with the fore wings. The knob, which is heavier than the rest of the organ, tends to keep the halteres beating in one plane. When the fly alters direction, whether voluntarily or otherwise, the haltere is twisted. The stem, which is richly endowed with sensilla, detects this movement, and the fly can respond accordingly.
Initiation of flight, for any reason, may involve the legs springing the insect into the air. Loss of tarsal contact with the ground causes neural firing of the direct flight muscles. In flies, flight activity originates in con- traction of a mid-leg muscle, which both propels the leg downwards (and the fly upwards) and simultaneously pulls the tergum downwards to inaugurate flight. The legs are also important when landing because there is no gradual braking by running forwards — all the shock is taken on the outstretched legs, endowed with pads, spines, and claws for adhesion.
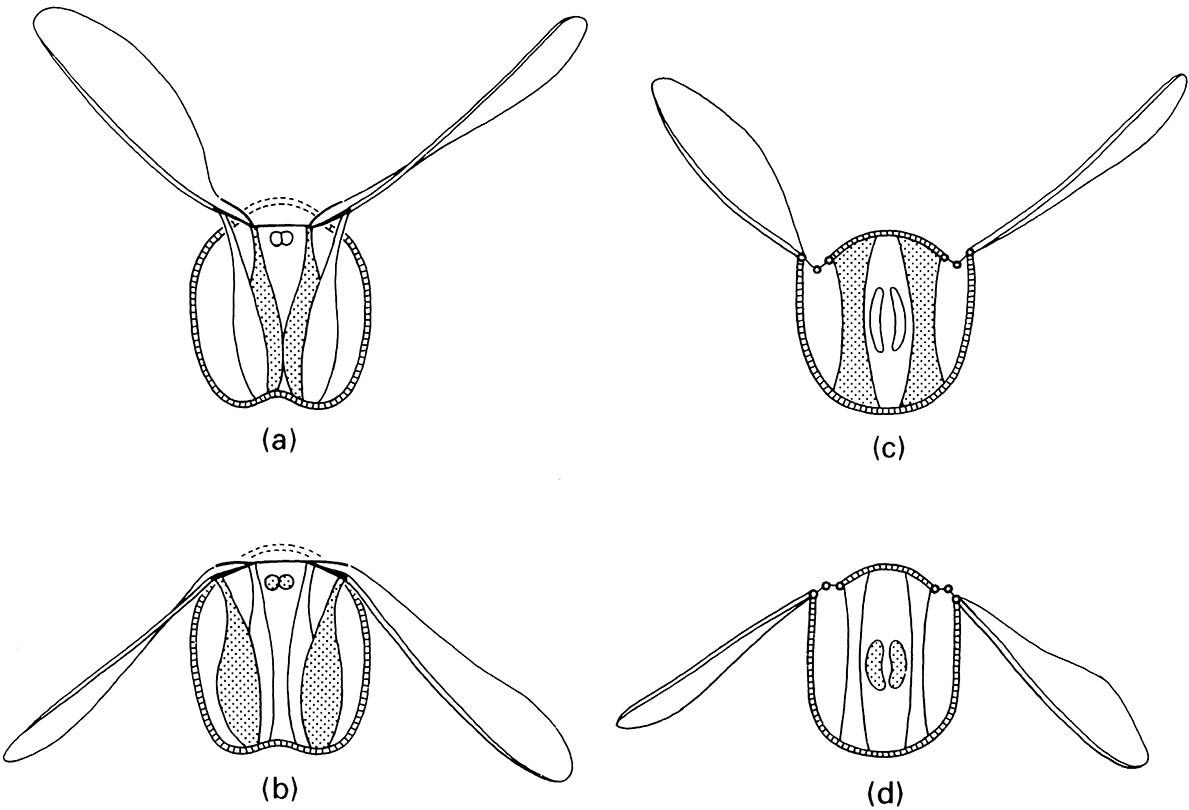
thorax during (c) upstroke and (d) downstroke of the wings. Stippled muscles are those contracting in each illustration. (After Blaney 1976)
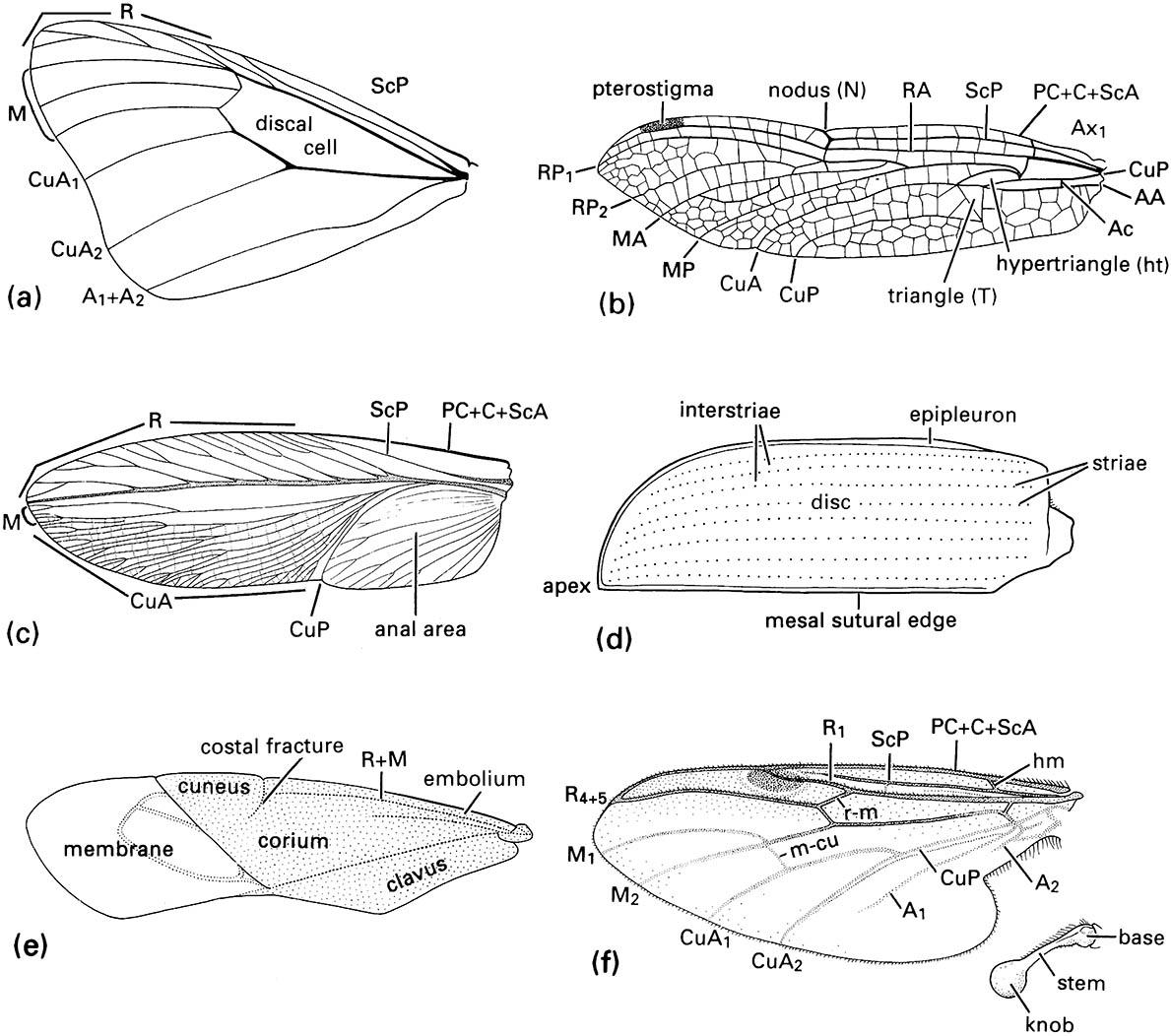
(a) fore wing of a butterfly of Danaus (Lepidoptera: Nymphalidae); (b) fore wing of a dragonfly of Urothemis (Odonata: Anisoptera: Libellulidae); (c) fore wing or tegmen of a cockroach of Periplaneta (Blattodea: Blattidae); (d) fore wing or elytron of a beetle of Anomala (Coleoptera: Scarabaeidae); (e) fore wing or hemelytron of a mirid bug (Hemiptera: Heteroptera: Miridae) showing three wing areas — the membrane, corium, and clavus; (f ) fore wing and haltere of a fly of Bibio (Diptera: Bibionidae) (after J.W.H. Trueman, unpublished. ((a-d) After Youdeowei 1977; (f) after McAlpine 1981)

