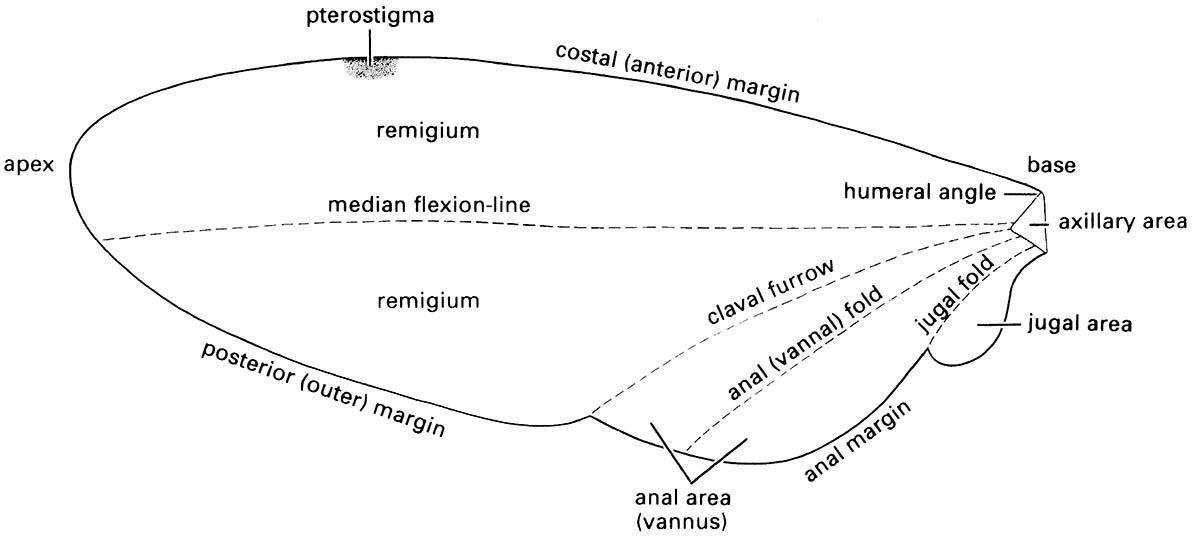
2.4.2. Wings
Wings are developed fully only in the adult, or exceptionally in the subimago, the penultimate stage of Ephemeroptera. Typically, functional wings are flap-like cuticular projections supported by tubular, sclerotized veins. The major veins are longitudinal, running from the wing base towards the tip, and are more concentrated at the anterior margin. Additional supporting cross-veins are transverse struts, which join the longitudinal veins to give a more complex structure. The major veins usually contain tracheae, blood vessels, and nerve fibers, with the intervening membranous areas comprising the closely appressed dorsal and ventral cuticular surfaces. Generally, the major veins are alternately “convex” and “concave” in relation to the surface plane of the wing, especially near the wing attachment; this configuration is described by plus (+) and minus (-) signs. Most veins lie in an anterior area of the wing called the remigium (Fig. 2.20), which, powered by the thoracic flight muscles, is responsible for most of the movements of flight. The area of wing posterior to the remigium sometimes is called the clavus; but more often two areas are recognized: an anterior anal area (or vannus) and a posterior jugal area. Wing areas are delimited and subdivided by fold-lines, along which the wing can be folded; and flexion-lines, at which the wing flexes during flight. The fundamental distinction between these two types of lines is often blurred, as fold-lines may permit some flexion and vice versa. The claval furrow (a flexion-line) and the jugal fold (or fold-line) are nearly constant in position in different insect groups, but the median flexion-line and the anal (or vannal) fold (or fold-line) form variable and unsatisfactory area boundaries. Wing folding may be very complicated; transverse folding occurs in the hind wings of Coleoptera and Dermaptera, and in some insects the enlarged anal area may be folded like a fan.
The fore and hind wings of insects in many orders are coupled together, which improves the aerodynamic efficiency of flight. The commonest coupling mechanism (seen clearly in Hymenoptera and some Trichoptera) is a row of small hooks, or hamuli, along the anterior margin of the hind wing that engages a fold along the posterior margin of the fore wing (hamulate coupling).
In some other insects (e.g. Mecoptera, Lepidoptera, and some Trichoptera), a jugal lobe of the fore wing overlaps the anterior hind wing ( jugate coupling), or the margins of the fore and hind wing overlap broadly (amplexiform coupling), or one or more hind-wing bristles (the frenulum) hook under a retaining structure (the retinaculum) on the fore wing (frenate coupling). The mechanics of flight are described in section 3.1.4 and the evolution of wings is covered in section 8.4.
All winged insects share the same basic wing venation comprising eight veins, named from anterior to posterior of the wing as: precosta (PC), costa (C), subcosta (Sc), radius (R), media (M), cubitus (Cu), anal (A), and jugal (J). Primitively, each vein has an anterior convex (+) sector (a branch with all of its subdivisions) and a posterior concave (-) sector. In almost all extant insects, the precosta is fused with the costa and the jugal vein is rarely apparent. The wing nomenclatural system presented in Fig. 2.21 is that of Kukalová-Peck and is based on detailed comparative studies of fossil and living insects. This system can be applied to the venation of all insect orders, although as yet it has not been widely applied because the various schemes devised for each insect order have a long history of use and there is a reluctance to discard familiar systems. Thus in most textbooks, the same vein may be referred to by different names in different insect orders because the structural homologies were not recognized correctly in early studies. For example, until 1991, the venational scheme for Coleoptera labeled the radius posterior (RP) as the media (M) and the media posterior (MP) as the cubitus (Cu). Correct interpretation of venational homologies is essential for phylogenetic studies and the establishment of a single, universally applied scheme is essential.
Cells are areas of the wing delimited by veins and may be open (extending to the wing margin) or closed (surrounded by veins). They are named usually according to the longitudinal veins or vein branches that they lie behind, except that certain cells are known by special names, such as the discal cell in Lepidoptera (Fig. 2.22a) and the triangle in Odonata (Fig. 2.22b). The pterostigma is an opaque or pigmented spot anteriorly near the apex of the wing (Figs. 2.20 & 2.22b).
Wing venation patterns are consistent within groups (especially families and orders) but often differ between groups and, together with folds or pleats, provide major features used in insect classification and identification. Relative to the basic scheme outlined above, venation may be greatly reduced by loss or postulated fusion of veins, or increased in complexity by numerous cross-veins or substantial terminal branching. Other features that may be diagnostic of the wings of different insect groups are pigment patterns and colors, hairs, and scales. Scales occur on the wings of Lepidoptera, many Trichoptera, and a few psocids (Psocoptera) and flies. Hairs consist of small microtrichia, either scattered or grouped, and larger macrotrichia, typically on the veins.
Usually two pairs of functional wings lie dorsolaterally as fore wings on the mesothorax and as hind wings on the metathorax; typically the wings are membranous and transparent. However, from this basic pattern are derived many other conditions, often involving variation in the relative size, shape, and degree of sclerotization of the fore and hind wings. Examples of fore-wing modification include the thickened, leathery fore wings of Blattodea, Dermaptera, and Orthoptera, which are called tegmina (singular: tegmen; Fig. 2.22c), the hardened fore wings of Coleoptera that form protective wing cases or elytra (singular: elytron; Fig. 2.22d & Plate 1.2), and the hemelytra (singular: hemelytron) of heteropteran Hemiptera with the basal part thickened and the apical part membranous (Fig. 2.22e). Typically, the heteropteran hemelytron is divided into three wing areas: the membrane, corium, and clavus. Sometimes the corium is divided further, with the embolium anterior to R + M, and the cuneus distal to a costal fracture. In Diptera the hind wings are modified as stabilizers (halteres) (Fig. 2.22f ) and do not function as wings, whereas in male Strepsiptera the fore wings form halteres and the hind wings are used in flight (Box 13.6). In male scale insects (see Plate 2.5, facing here) the fore wings have highly reduced venation and the hind wings form hamulohalteres (different in structure to the halteres) or are lost completely.
Small insects confront different aerodynamic challenges compared with larger insects and their wing area often is expanded to aid wind dispersal. Thrips (Thysanoptera), for example, have very slender wings but have a fringe of long setae or cilia to extend the wing area (Box 11.7). In termites (Isoptera) and ants (Hymenoptera: Formicidae) the winged reproductives, or alates, have large deciduous wings that are shed after the nuptial flight. Some insects are wingless, or apterous, either primitively as in silverfish (Zygentoma) and bristletails (Archaeognatha), which diverged from other insect lineages prior to the origin of wings, or secondarily as in all lice (Phthiraptera) and fleas (Siphonaptera), which evolved from winged ancestors. Secondary partial wing reduction occurs in a number of short-winged, or brachypterous, insects.
In all winged insects (Pterygota), a triangular area at the wing base, the axillary area (Fig. 2.20), contains the movable articular sclerites via which the wing articulates on the thorax. These sclerites are derived, by reduction and fusion, from a band of articular sclerites in the ancestral wing. Three different types of wing articulation among living Pterygota result from unique patterns of fusion and reduction of the articular sclerites. In Neoptera (all living winged insects except the Ephemeroptera and Odonata), the articular sclerites consist of the humeral plate, the tegula, and usually three, rarely four, axillary sclerites (1Ax, 2Ax, 3Ax, and 4Ax) (Fig. 2.21). The Ephemeroptera and Odonata each has a different configuration of these sclerites compared with the Neoptera (literally meaning “new wing”). Odonate and ephemeropteran adults cannot fold their wings back along the abdomen as can neopterans. In Neoptera, the wing articulates via the articular sclerites with the anterior and posterior wing processes dorsally, and ventrally with the pleural wing processes and two small pleural sclerites (the basalare and subalare) (Fig. 2.18).
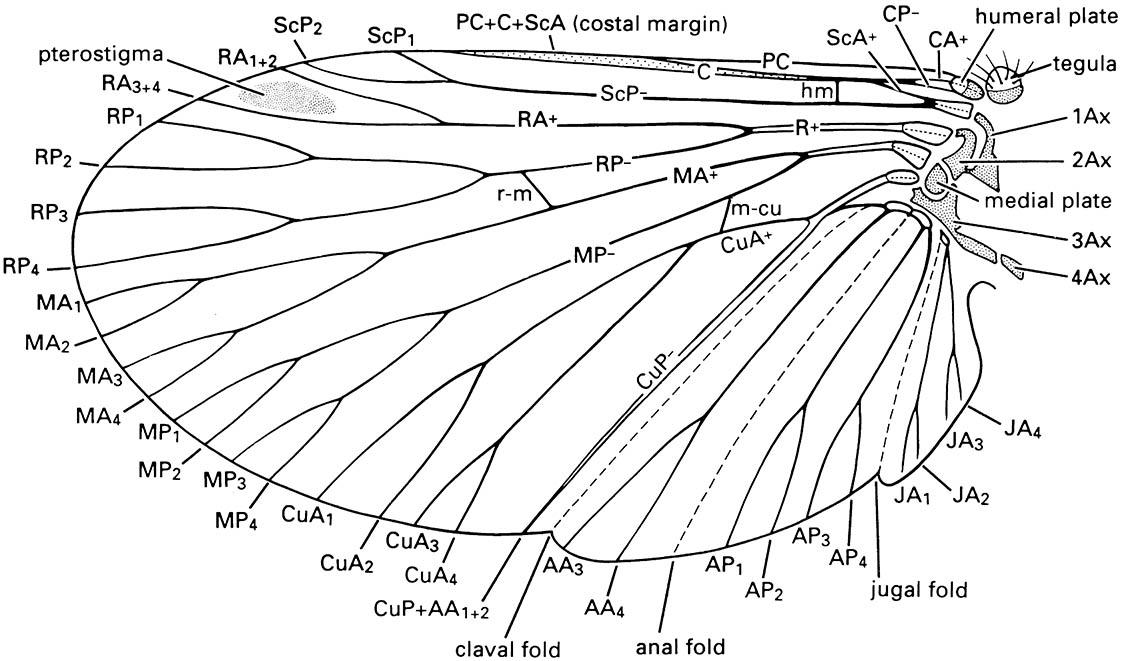
Notation as follows: AA, anal anterior; AP, anal posterior; Ax, axillary sclerite; C, costa; CA, costa anterior; CP, costa posterior; CuA, cubitus anterior; CuP, cubitus posterior; hm, humeral vein; JA, jugal anterior; MA, media anterior; m-cu, cross-vein between medial and cubital areas; MP, media posterior; PC, precosta; R, radius; RA, radius anterior; r-m, cross-vein between radial and median areas; RP, radius posterior; ScA, subcosta anterior; ScP, subcosta posterior. Branches of the anterior and posterior sector of each vein are numbered, e.g. CuA 1-4. (After CSIRO 1991)
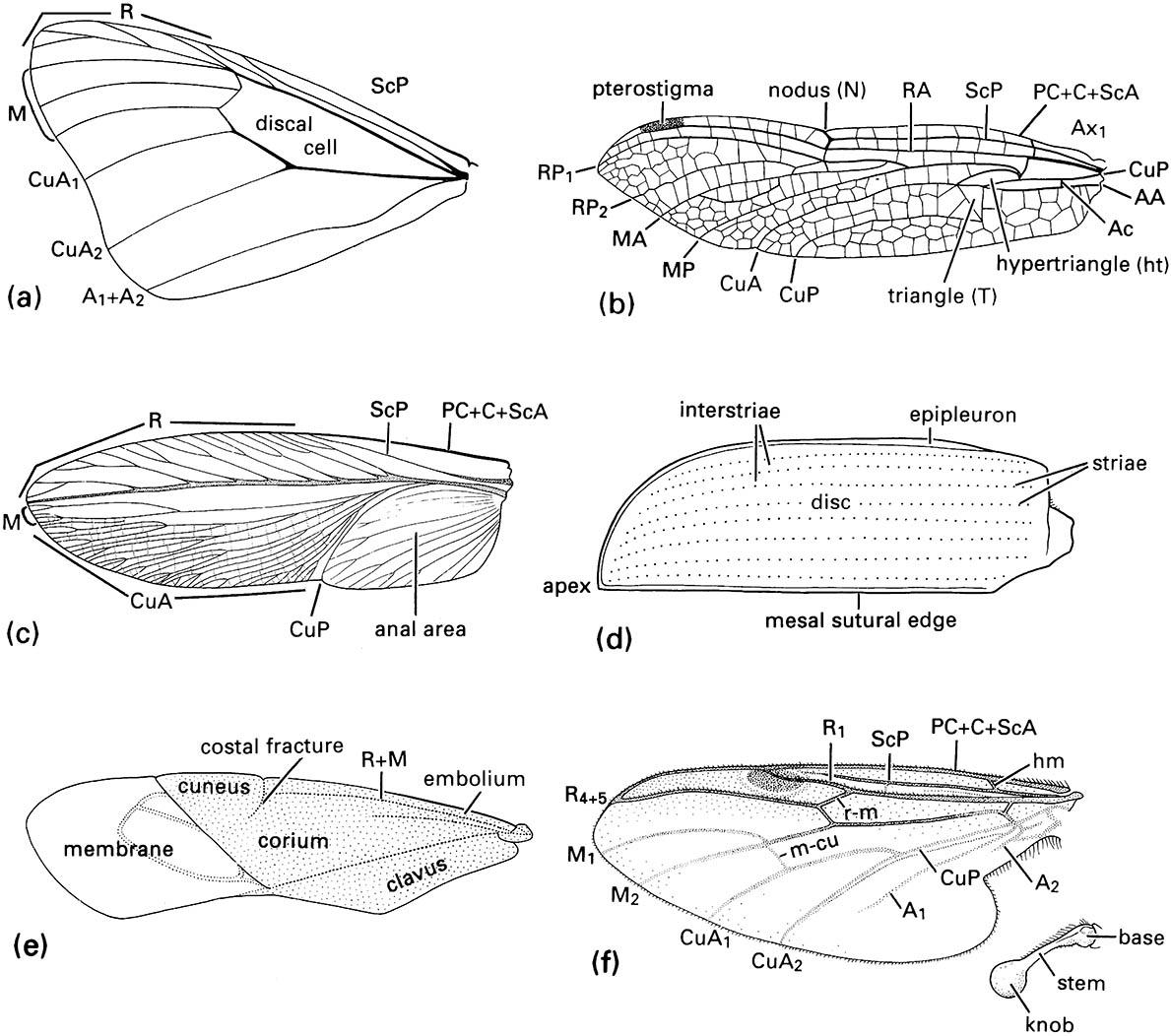
(a) fore wing of a butterfly of Danaus (Lepidoptera: Nymphalidae); (b) fore wing of a dragonfly of Urothemis (Odonata: Anisoptera: Libellulidae); (c) fore wing or tegmen of a cockroach of Periplaneta (Blattodea: Blattidae); (d) fore wing or elytron of a beetle of Anomala (Coleoptera: Scarabaeidae); (e) fore wing or hemelytron of a mirid bug (Hemiptera: Heteroptera: Miridae) showing three wing areas — the membrane, corium, and clavus; (f ) fore wing and haltere of a fly of Bibio (Diptera: Bibionidae) (after J.W.H. Trueman, unpublished. ((a-d) After Youdeowei 1977; (f) after McAlpine 1981)
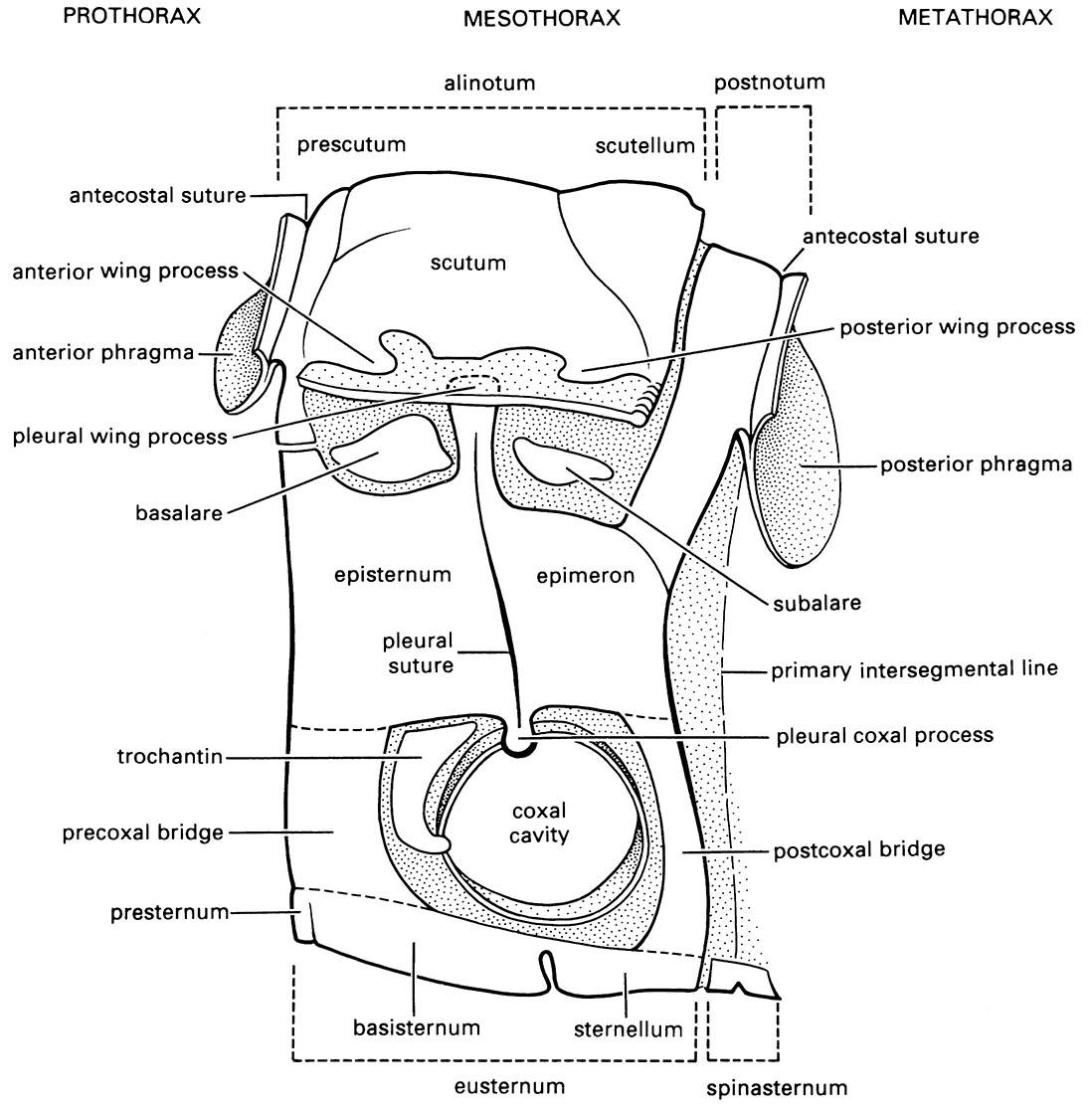
(After Snodgrass 1935)

