6.2.1. Embryonic phase
The egg stage begins as soon as the female deposits the mature egg. For practical reasons, the age of an egg is estimated from the time of its deposition even though the egg existed before oviposition. The beginning of the egg stage, however, need not mark the commencement of an individual insect’s ontogeny, which actually begins when embryonic development within the egg is triggered by activation. This trigger usually results from fertilization in sexually reproducing insects, but in parthenogenetic species appears to be induced by various events at oviposition, including the entry of oxygen to the egg or mechanical distortion.
Following activation of the insect egg cell, the zygote nucleus subdivides by mitotic division to produce many daughter nuclei, giving rise to a syncytium. These nuclei and their surrounding cytoplasm, called cleavage energids, migrate to the egg periphery where the membrane infolds leading to cellularization of the superficial layer to form the one-cell thick blastoderm. This distinctive superficial cleavage during early embryogenesis in insects is the result of the large amount of yolk in the egg. The blastoderm usually gives rise to all the cells of the larval body, whereas the central yolky part of the egg provides the nutrition for the developing embryo and will be used up by the time of eclosion, or emergence from the egg.
Regional differentiation of the blastoderm leads to the formation of the germ anlage or germ disc (Fig.6.5a), which is the first sign of the developing embryo, whereas the remainder of the blastoderm becomes a thin membrane, the serosa, or embryonic cover. Next, the germ anlage develops an infolding in a process called gastrulation (Fig. 6.5b) and sinks into the yolk, forming a two-layered embryo containing the amniotic cavity (Fig. 6.5c). After gastrulation, the germ anlage becomes the germ band, which externally is characterized by segmental organization (commencing in Fig. 6.5d with the formation of the protocephalon). The germ band essentially forms the ventral regions of the future body, which progressively differentiates with the head, body segments, and appendages becoming increasingly well defined (Fig. 6.5e-g). At this time the embryo undergoes movement called katatrepsis which brings it into its final position in the egg. Later, near the end of embryogenesis (Fig. 6.5h,i), the edges of the germ band grow over the remaining yolk and fuse mid-dorsally to form the lateral and dorsal parts of the insect — a process called dorsal closure.
In the well-studied Drosophila, the complete embryo is large, and becomes segmented at the cellularization stage, termed “long germ” (as in all studied Diptera, Coleoptera, and Hymenoptera). In contrast, in “short-germ” insects (phylogenetically earlier branching taxa, including locusts) the embryo derives from only a small region of the blastoderm and the posterior segments are added post-cellularization, during subsequent growth. In the developing long-germ embryo, the syncytial phase is followed by cell membrane intrusion to form the blastoderm phase.
Functional specialization of cells and tissues occurs during the latter period of embryonic development, so that by the time of hatching (Fig. 6.5j) the embryo is a tiny proto-insect crammed into an eggshell. In ametabolous and hemimetabolous insects, this stage may be recognized as a pronymph — a special hatching stage (section 8.5). Molecular developmental processes involved in organizing the polarity and differentiation of areas of the body, including segmentation, are reviewed in Box 6.1.
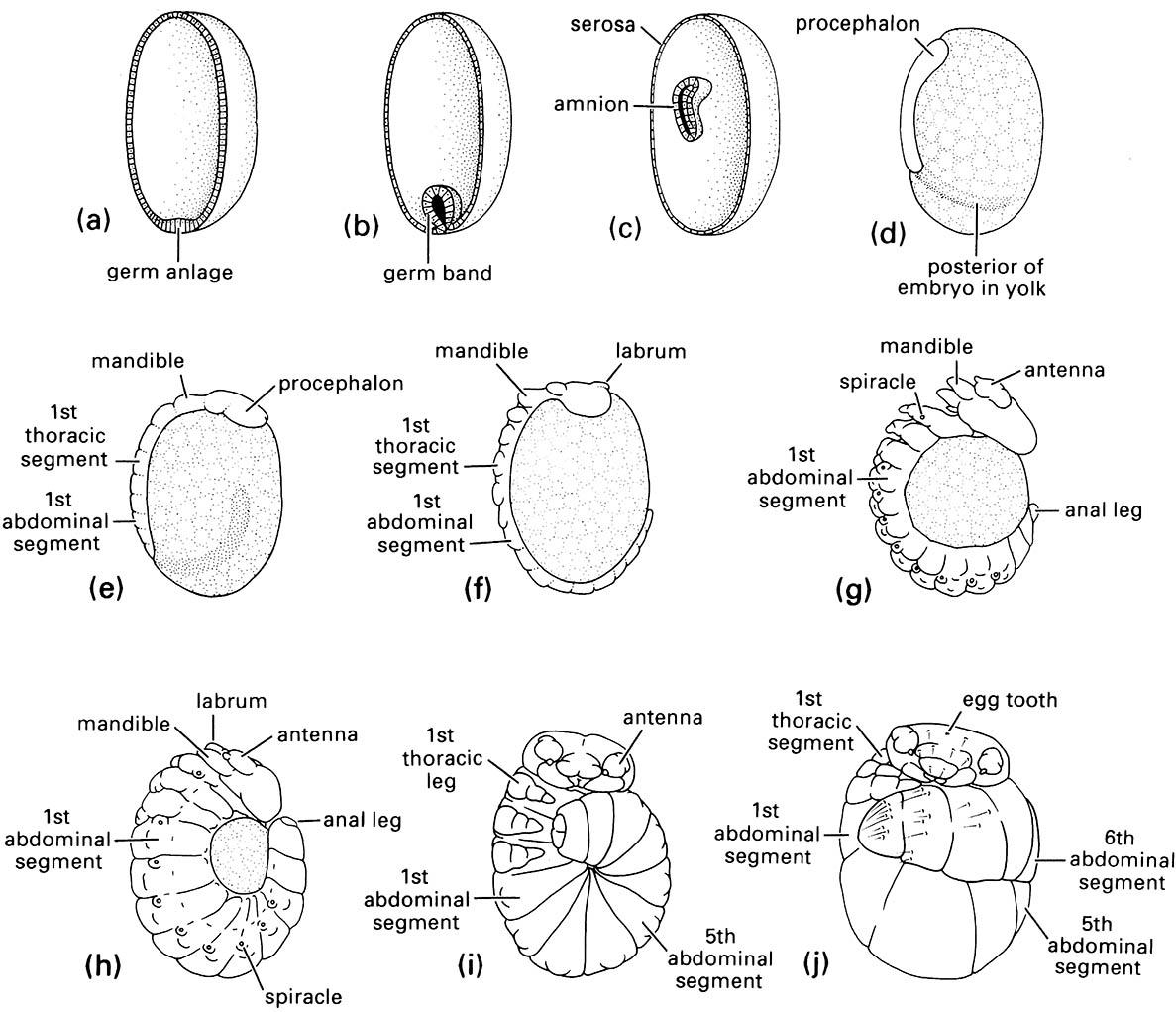
(a-c) schematic drawings of egg halves from which yolk has been removed to show position of embryo; (d-j) gross morphology of developing embryos at various ages. Age from oviposition: (a) 32 h; (b) 2 days; (c) 7 days; (d) 12 days; (e) 16 days; (f ) 19 days; (g) 23 days; (h) 25 days; (i) 25-26 days; (j) full grown at 32 days. (After Suzuki 1985)

