6.10.1. Temperature
Most insects are poikilothermic, that is with body temperature more or less directly varying with environmental temperature, thus heat is the force driving the rate of growth and development when food is unlimited. A rise in temperature, within a favorable range, will speed up the metabolism of an insect and consequently increase its rate of development. Each species and each stage in the life history may develop at its own rate in relation to temperature. Thus, physiological time, a measure of the amount of heat required over time for an insect to complete development or a stage of development, is more meaningful as a measure of development time than age in calendar time. Knowledge of temperature—development relationships and the use of physiological time allow comparison of the life cycles and/or fecundity of pest species in the same system (Fig. 6.12), and prediction of the larval feeding periods, generation length, and time of adult emergence under variable temperature conditions that exist in the field. Such predictions are especially important for pest insects, as control measures must be timed carefully to be effective.
Physiological time is the cumulative product of total development time (in hours or days) multiplied by the temperature (in degrees) above the developmental (or growth) threshold, or the temperature below which no development occurs. Thus, physiological time is commonly expressed as day-degrees (D°) or hour- degrees (h°). Normally, physiological time is estimated for a species by rearing a number of individuals of the life-history stage(s) of interest under different constant temperatures in several identical growth cabinets. The developmental threshold is estimated by the linear regression x-axis method, as outlined in Box 6.2, although more accurate threshold estimates can be obtained by more time-consuming methods.
In practice, the application of laboratory-estimated physiological time to natural populations may be complicated by several factors. Under fluctuating temperatures, especially if the insects experience extremes, growth may be retarded or accelerated compared with the same number of day-degrees under constant temperatures. Furthermore, the temperatures actually experienced by the insects, in their often sheltered microhabitats on plants or in soil or litter, may be several degrees different from the temperatures recorded at a meteorological station even just a few meters away. Insects may select microhabitats that ameliorate cold night conditions or reduce or increase daytime heat. Thus, predictions of insect life-cycle events based on extrapolation from laboratory to field temperature records may be inaccurate. For these reasons, the laboratory estimates of physiological time should be corroborated by calculating the hour-degrees or day-degrees required for development under more natural conditions, but using the laboratory-estimated develop- mental threshold, as follows.
- Place newly laid eggs or newly hatched larvae in their appropriate field habitat and record temperature each hour (or calculate a daily average — a less accurate method).
- Estimate the time for completion of each instar by discarding all temperature readings below the developmental threshold of the instar and subtracting the developmental threshold from all other readings to determine the effective temperature for each hour (or simply subtract the development threshold temperature from the daily average temperature). Sum the degrees of effective temperature for each hour from the beginning to the end of the stadium. This procedure is called thermal summation.
- Compare the field-estimated number of hour-degrees (or day-degrees) for each instar with that predicted from the laboratory data. If there are discrepancies, then microhabitat and/or fluctuating temperatures may be influencing insect development or the developmental zero read from the graph may be a poor estimate of the developmental threshold.
Another problem with laboratory estimation of physiological time is that insect populations maintained for lengthy periods under laboratory conditions frequently undergo acclimation to constant conditions or even genetic change in response to the altered environment or as a result of population reductions that produce genetic “bottle-necks”. Therefore, insects maintained in rearing cages may exhibit different temperature—development relationships from individuals of the same species in wild populations.
For all of the above reasons any formula or model that purports to predict insect response to environmental conditions must be tested carefully for its fit with natural population responses.
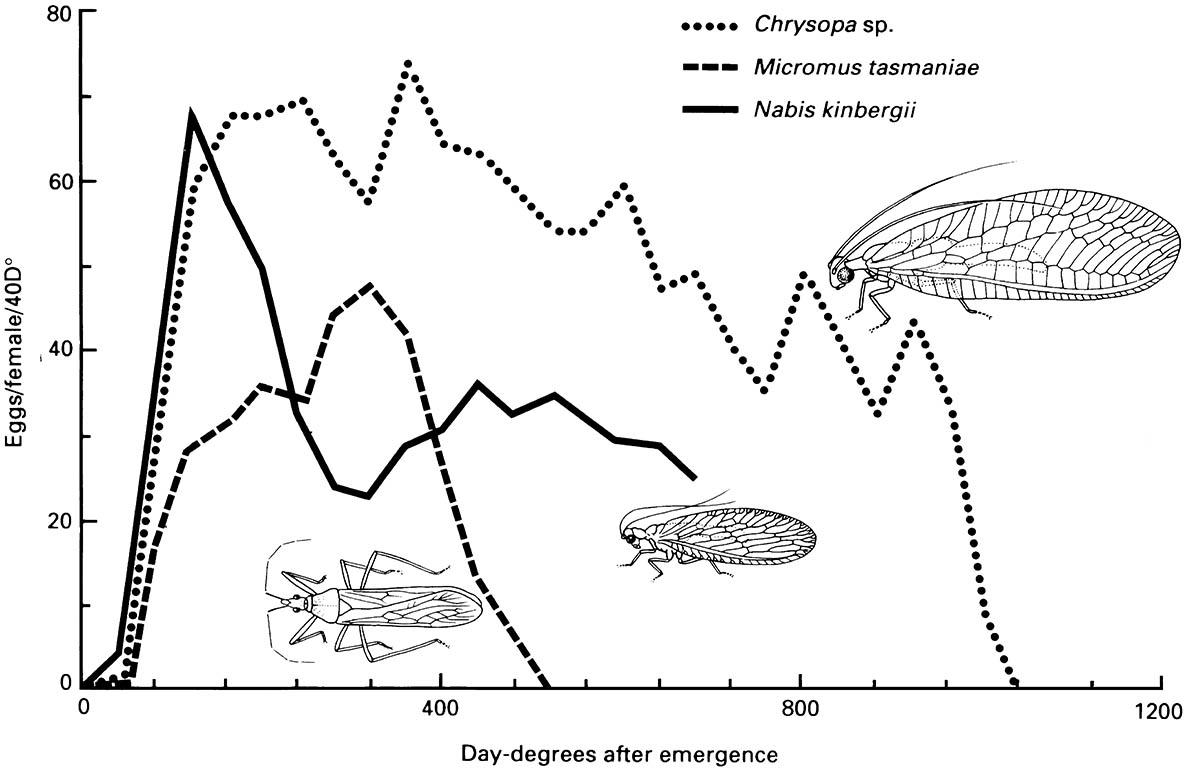
(After Samson & Blood 1979)

