8.5. Evolution of metamorphosis
As we have seen earlier, the evolution of metamorphosis — which allows larval immature stages to be separated ecologically from the adult stage to avoid competition — seems to have been an important factor encouraging diversification.
Although how holometaboly (with larval juvenile instars highly differentiated from adults by metamorphosis) evolved from incremental hemimetaboly has been debated, strong support for one candidate hypothesis is now available. This involves a proposal that a pronymph (hatchling or pre-hatching stage, distinct from subsequent nymphal stages) is the evolutionary precursor to the holometabolous larva, and the holometabolous pupa is the sole nymphal stage.
In ametabolous taxa, which form the earliest branches in the hexapodan phylogeny, at each molt the subsequent instar is a larger version of the previous, and development is linear, progressive, and continuous. Even early flying insects, such as the Palaeodictyoptera (Fig. 8.2), in all stages (sizes) of fossil nymphs had pro- portionally scaled winglets, and thus were ametabolous. A distinctive earliest developmental stage, the pronymph, forms an exception to the proportionality of nymphal development. The pronymph stage, which feeds only on yolk reserves, can survive independently and move for some days after hatching. Hemimetabolous insects, which differ from ametabolous taxa in that the adult instar with fully formed genitalia and wings undergoes no further molting, also have a recognizably distinct pronymph.
The body proportions of the pronymph differ from those of subsequent nymphal stages, perhaps con- strained by confinement inside the egg and by the need to assist in hatching (if indeed this stage is that which hatches). Clearly, the pterygote pronymph is not just a highly miniaturized first-instar nymph. In certain orders (Blattodea, Phthiraptera, Hemiptera) the hatch- ling may be a pharate first-instar nymph, inside the pronymphal cuticle. At hatching the nymph emerges from the egg, since the first molt occurs concurrently with eclosion. In Odonata and Orthoptera the hatchling is the actual pronymph which can undertake limited, often specialized, post-hatching movement to locate a potential nymphal development site before molting to the first true nymph.
The larval stages of Holometabola are theorized to be homologous to this pronymphal stage, and the hemimetabolous nymphal stages are contracted into the holometabolous pupa, which is the only nymphal stage. Supporting evidence for this view comes from recognition of differences between pronymphal, nymphal, and larval cuticle, the timing of different cuticle formations relative to embryogenetic stages (katatrepsis — adoption of the final position in egg — and dorsal closure; see section 6.2.1), and interruption of neuroblast-induced neuron production during larval stages, which resumes in the nymph.
The mechanism that could cause such dramatic changes in development is heterochrony — alteration in the timing of activation of different controls involved in developmental cascades (Box 6.1). Metamorphosis is controlled by the interplay between neuropeptides, ecdysteroids, and especially juvenile hormone ( JH) titers, as seen in section 6.3. The balance between controlling factors commences in the egg, and continues throughout development: subtle differences in timing of events lead to very different outcomes. Earlier appearance of elevated JH in the embryo prevents maturity of some aspects of the nymph, leading to development of a prolarva, which then is maintained in larval form by continued high JH which suppresses maturation. Pupation (entry to the nymphal stage) takes place when JH is reduced, and maturation then requires increased JH. Holometabolous development occurs because JH remains high, with the JH-free period delayed until the end of immature growth (metamorphosis). This contrasts with hemimetaboly, in which postembryonic, continuous low JH exposure allows nymphal development to progress evenly towards the adult form.
In early-branching (“primitive”) Holometabola, JH prevents any precocious production of adult features in the larva until the pupa. However, in later-branching orders and more derived families some adult features can escape suppression by JH and may commence development in early larval instars. Such features include wings, legs, antennae, eyes, and genitalia: their early expression is seen in groups of primordial cells that become imaginal discs — differentiated already for their final adult function (section 6.2, Fig. 6.4). With scope to vary the onset of differentiation of each adult organ in the larvae, great variation and flexibility in life-cycle evolution is permitted, including capacity to greatly shorten them.
Timing of developmental control is evident also in the basic larval shape, especially the variety of larval leg forms shown in Fig. 6.6. Onset of JH expression can retard development of the pronymph/prolarva at any stage in leg expression, from apodous (no expression) to essentially fully developed. Such legs, although termed prolegs in larvae (section 2.4.1), have innervation similar to, but less developed than, that found in adult legs. Evidently, immature and imaginal legs are homologous — since adult leg imaginal discs develop within the prolegs, no matter how well developed the prolegs are.
The major unanswered question in this view of the evolution of holometaboly is that if larval evolution results from a protracted equivalent of the pronymphal stage, how did the pronymph become able to feed? Although crustacean pronymphs (e.g. the nauplius stage of decapods) feed, the equivalent stage in extant hexapods apparently cannot do so. The pronymph has a short post-hatching existence, but if it finds itself in a suitable microhabitat by female oviposition-site selection or its own limited ability to search, and could feed, then there would be a tendency to select for this ability in ensuing instars. Selection is seen as continuing, because decoupling of larval and adult food resources reduces competition between juvenile and adult for food, thus separating resources used in growth from those for reproduction. The evident success of Holometabola derives not least from this segregation.
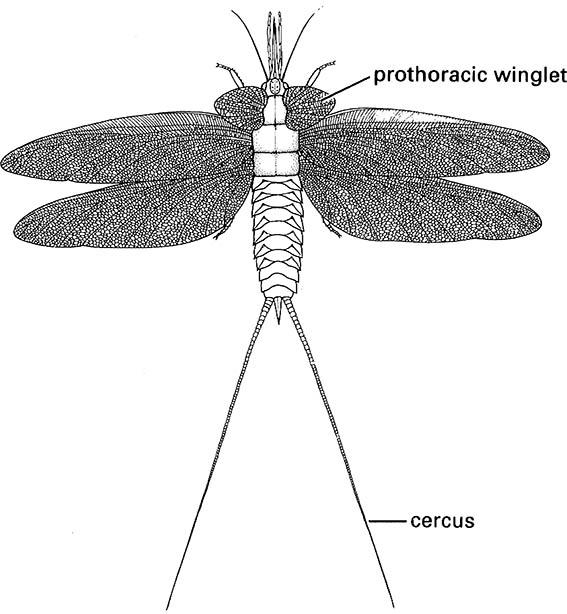
(After Kukalová 1970)
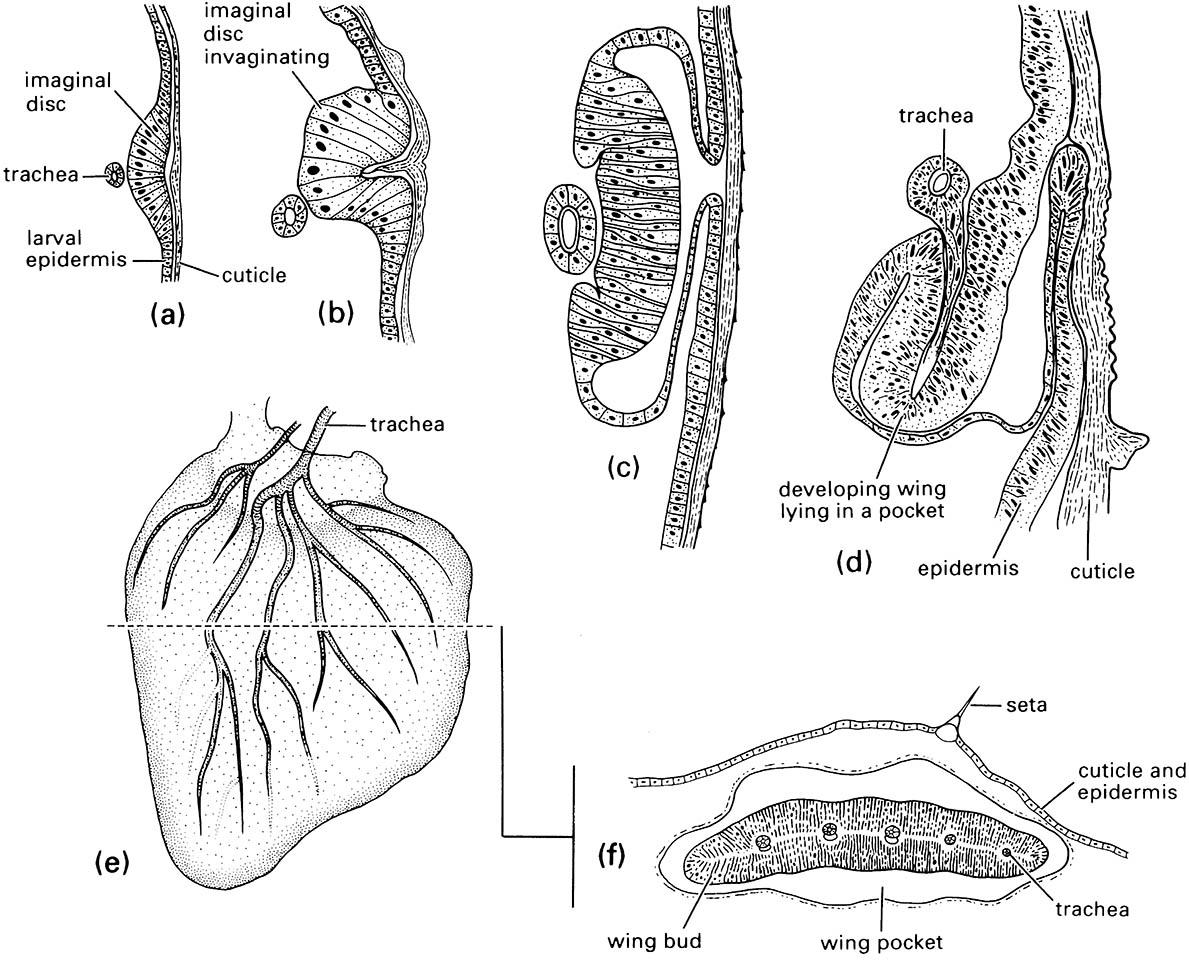
A wing imaginal disc in an (a) first-instar larva, (b) second-instar larva, (c) third-instar larva, and (d) fourth-instar larva; (e) the wing bud as it appears if dissected out of the wing pocket or (f ) cut in cross-section in a fifth-instar larva. ((a-e) After Mercer 1900)
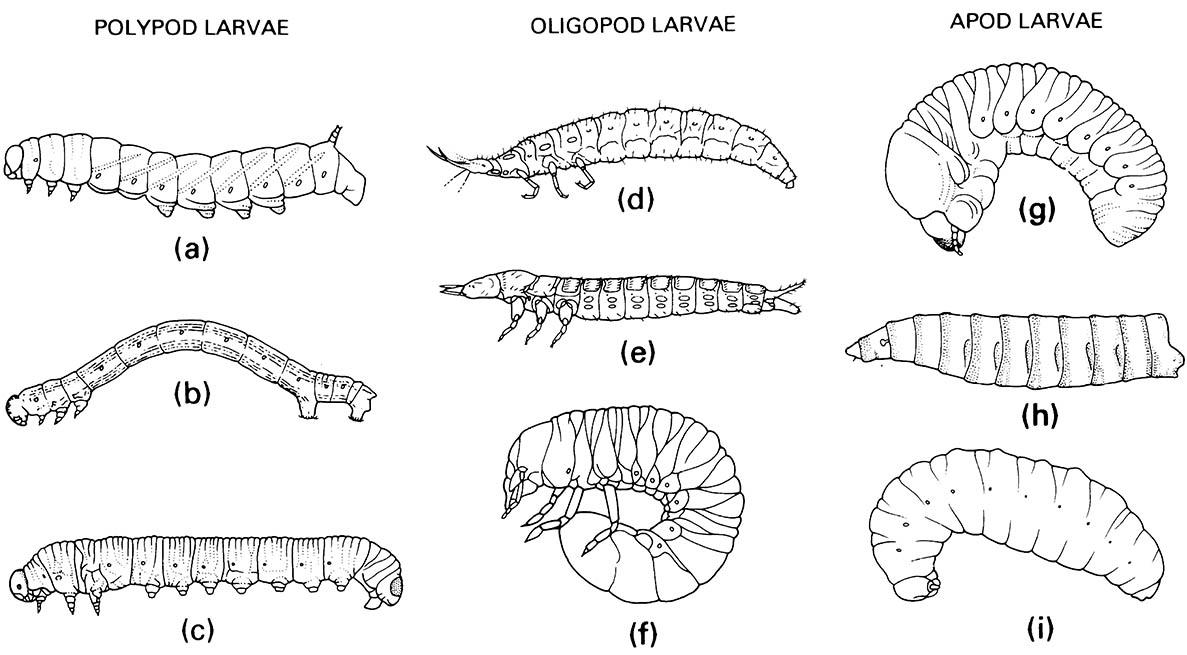
(a) Lepidoptera: Sphingidae; (b) Lepidoptera: Geometridae; (c) Hymenoptera: Diprionidae. Oligopod larvae: (d) Neuroptera: Osmylidae; (e) Coleoptera: Carabidae; (f ) Coleoptera: Scarabaeidae. Apod larvae: (g) Coleoptera: Scolytidae; (h) Diptera: Calliphoridae; (i) Hymenoptera: Vespidae. ((a, e-g) After Chu 1949; (b, c) after Borror et al. 1989; (h) after Ferrar 1987; (i) after CSIRO 1970)

