6.2.3. Metamorphosis
All pterygote insects undergo varying degrees of trans- formation from the immature to the adult phase of their life history. Some exopterygotes, such as cockroaches, show only slight morphological changes during post-embryonic development, whereas the body is largely reconstructed at metamorphosis in many endopterygotes. Only the Holometabola (= Endopterygota) have a metamorphosis involving a pupal stadium, during which adult structures are elaborated from larval structures. Alterations in body shape, which are the essence of metamorphosis, are brought about by differential growth of various body parts. Organs that will function in the adult but that were undeveloped in the larva grow at a faster rate than the body average. The accelerated growth of wing pads is the most obvious example, but legs, genitalia, gonads, and other internal organs may increase in size and complexity to a considerable extent.
The onset of metamorphosis generally is associated with the attainment of a certain body size, which is thought to program the brain for metamorphosis, resulting in altered hormone levels. Metamorphosis in most studied beetles, however, shows considerable independence from the influence of the brain, especially during the pupal instar. In most insects, a reduction in the amount of circulating juvenile hormone (as a result of reduction of corpora allata activity) is essential to the initiation of metamorphosis. (The physiological events are described in section 6.3.)
The molt into the pupal instar is called pupation, or the larval—pupal molt. Many insects survive conditions unfavorable for development in the “resting”, non-feeding pupal stage, but often what appears to be a pupa is actually a fully developed adult within the pupal cuticle, referred to as a pharate (cloaked) adult. Typically, a protective cell or cocoon surrounds the pupa and then, prior to emergence, the pharate adult; only certain Coleoptera, Diptera, Lepidoptera, and Hymenoptera have unprotected pupae.
Several pupal types (Fig. 6.7) are recognized and these appear to have arisen convergently in different orders. Most pupae are exarate (Fig. 6.7a-d) — their appendages (e.g. legs, wings, mouthparts, and antennae) are not closely appressed to the body (see Plate 3.2, facing here); the remaining pupae are obtect (Fig. 6.7g-j) — their appendages are cemented to the body and the cuticle is often heavily sclerotized (as in almost all Lepidoptera). Exarate pupae can have articulated mandibles (decticous), that the pharate adult uses to cut through the cocoon, or the mandibles can be non-articulated (adecticous), in which case the adult usually first sheds the pupal cuticle and then uses its mandibles and legs to escape the cocoon or cell. In some cyclorrhaphous Diptera (the Schizophora) the adecticous exarate pupa is enclosed in a puparium (Fig. 6.7e,f ) — the sclerotized cuticle of the last larval instar. Escape from the puparium is facilitated by eversion of a membranous sac on the head of the emerging adult, the ptilinum. Insects with obtect pupae may lack a cocoon, as in coccinellid beetles and most nematocerous and orthorrhaphous Diptera. If a cocoon is present, as in most Lepidoptera, emergence from the cocoon is either by the pupa using backwardly directed abdominal spines or a projection on the head, or an adult emerges from the pupal cuticle before escaping the cocoon, sometimes helped by a fluid that dissolves the silk.
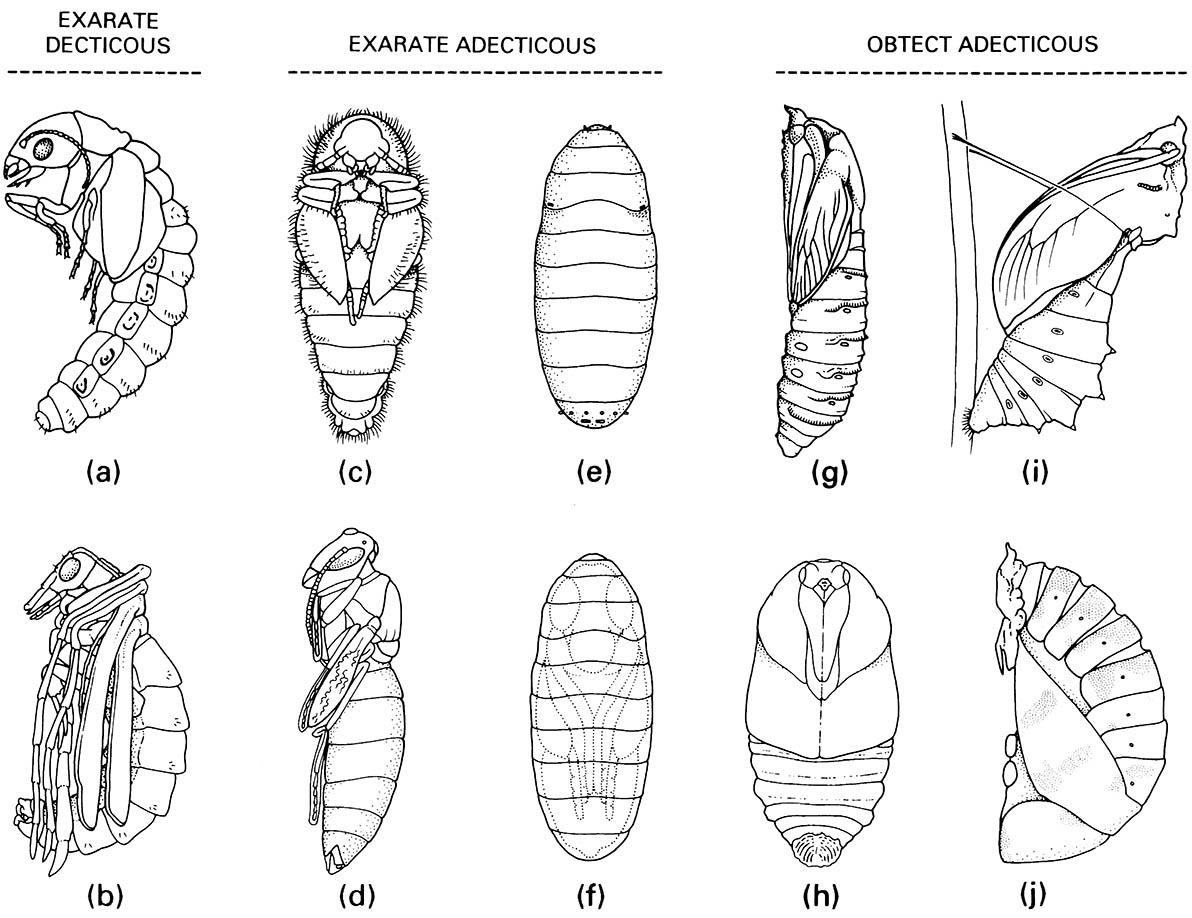
Exarate decticous pupae: (a) Megaloptera: Sialidae; (b) Mecoptera: Bittacidae. Exarate adecticous pupae: (c) Coleoptera: Dermestidae; (d) Hymenoptera: Vespidae; (e,f ) Diptera: Calliphoridae, puparium and pupa within. Obtect adecticous pupae: (g) Lepidoptera: Cossidae; (h) Lepidoptera: Saturniidae; (i) Lepidoptera: Papilionidae, chrysa lis; (j) Coleoptera: Coccinellidae. ((a) After Evans 1978; (b, c, e, g) after CSIRO 1970; (d) after Chu 1949; (h) after Common 1990; (i) after Common & Waterhouse 1972; (j) after Palmer 1914)

