6.2.4. Imaginal or adult phase
Except for the mayflies, insects do not molt again once the adult phase is reached. The adult, or imaginal, stage has a reproductive role and is often the dispersive stage in insects with relatively sedentary larvae. After the imago emerges from the cuticle of the previous instar (eclosion), it may be reproductively competent almost immediately or there may be a period of maturation in readiness for sperm transfer or oviposition. Depending on species and food availability, there are from one to several reproductive cycles in the adult stadium. The adults of certain species, such as some mayflies, midges, and male scale insects, are very short-lived. These insects have reduced or no mouthparts and fly for only a few hours or at the most a day or two — they simply mate and die. Most adult insects live at least a few weeks, often a few months and sometimes for several years; termite reproductives and queen ants and bees are particularly long-lived.
Adult life begins at eclosion from the pupal cuticle. Metamorphosis, however, may have been complete for some hours, days, or weeks previously and the pharate adult may have rested in the pupal cuticle until the appropriate environmental trigger for emergence. Changes in temperature or light and perhaps chemical signals may synchronize adult emergence in most species.
Hormonal control of emergence has been studied most comprehensively in Lepidoptera, especially in the tobacco hornworm, Manduca sexta (Lepidoptera: Sphingidae), notably by James Truman, Lynn Riddiford, and colleagues. The description of the following events at eclosion are based largely on M. sexta but are believed to be similar in other insects and at other molts. At least five hormones are involved in eclosion (see also section 6.3). A few days prior to eclosion the ecdysteroid level declines, and a series of physiological and behavioral events are initiated in preparation for ecdysis, including the release of two neuropeptides. Ecdysis triggering hormone (ETH), from epitracheal glands called Inka cells, and eclosion hormone (EH), from neurosecretory cells in the brain, act in concert to trigger pre-eclosion behavior, such as seeking a site suitable for ecdysis and movements to aid later extrication from the old cuticle. ETH is released first and ETH and EH stimulate each other’s release, forming a positive feedback loop. The build-up of EH also releases crustacean cardioactive peptide (CCAP) from cells in the ventral nerve cord. CCAP switches off pre-eclosion behavior and switches on eclosion behavior, such as abdominal contraction and wing-base movements, and accelerates heartbeat. EH appears also to permit the release of further neurohormones — bursicon and cardiopeptides — that are involved in wing expansion after ecdysis. The cardiopeptides stimulate the heart, facilitating movement of hemolymph into the thorax and thus into the wings. Bursicon induces a brief increase in cuticle plasticity to permit wing expansion, followed by sclerotization of the cuticle in its expanded form.
The newly emerged, or teneral, adult has soft cuticle, which permits expansion of the body surface by swallowing air, by taking air into the tracheal sacs, and by locally increasing hemolymph pressure by muscular activity. The wings normally hang down (Fig. 6.8; see also Plate 3.4), which aids their inflation. Pigment deposition in the cuticle and epidermal cells occurs just before or after emergence and is either linked to, or followed by, sclerotization of the body cuticle under the influence of the neurohormone bursicon.
Following emergence from the pupal cuticle, many holometabolous insects void a fecal fluid called the meconium. This represents the metabolic wastes that have accumulated during the pupal stadium. Sometimes the teneral adult retains the meconium in the rectum until sclerotization is complete, thus aiding increase in body size.
Reproduction is the main function of adult life and the length of the imaginal stadium, at least in the female, is related to the duration of egg production. Reproduction is discussed in detail in Chapter 5. Senescence correlates with termination of reproduction and death may be predetermined in the ontogeny of an insect. Females may die after egg deposition and males may die after mating. An extended post-reproductive life is important in distasteful, aposematic insects to allow predators to learn the distastefulness of the prey at a developmental period when prey individuals are expendable (section 14.4).
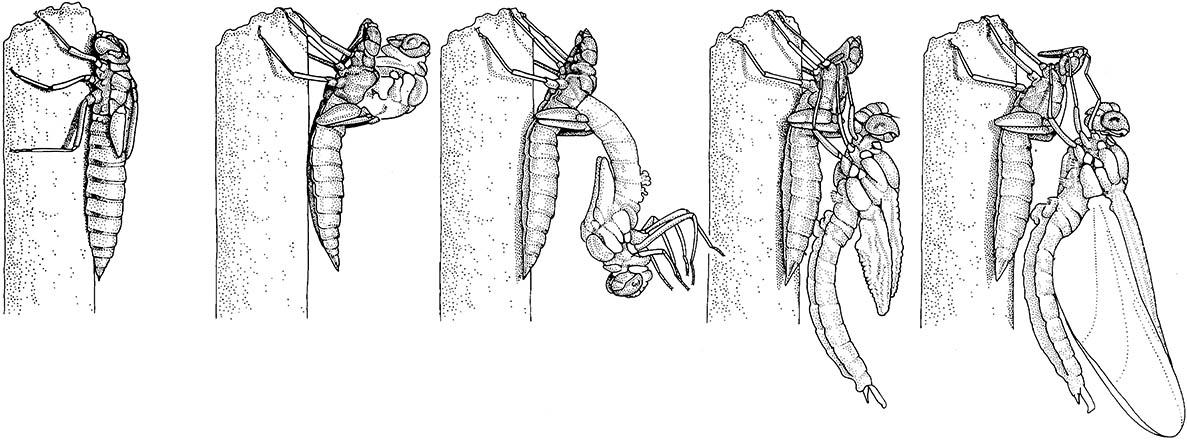
The final-instar nymph climbs out of the water prior to the shedding of its cuticle. The old cuticle splits mid-dorsally, the teneral adult frees itself, swallows air and must wait many hours for its wings to expand and dry. (After Blaney 1976)

