4.3.2. Semiochemicals: pheromones
Many insect behaviors rely on the sense of smell. Chemical odors, termed semiochemicals (from semion — signal), are especially important in both interspecific and intraspecific communication. The latter is particularly highly developed in insects, and involves the use of chemicals called pheromones. When recognized first in the 1950s, pheromones were defined as: substances that are secreted to the outside by one individual and received by a second individual of the same species in which they release a specific reaction, for example a definite behavior or developmental process. This definition remains valid today, despite the discovery of a hidden complexity of pheromone cocktails.
Pheromones are predominantly volatile but some- times are liquid contact chemicals. All are produced by exocrine glands (those that secrete to the outside of the body) derived from epidermal cells. The scent organs may be located almost anywhere on the body. Thus, sexual scent glands on female Lepidoptera lie in eversible sacs or pouches between the eighth and ninth abdominal segments; the organs are mandibular in the female honey bee, but are located on the swollen hind tibiae of female aphids, and within the midgut and genitalia in cockroaches.
Classification of pheromones by chemical structure reveals that many naturally occurring compounds (such as host odors) and preexisting metabolites (such as cuticular waxes) have been co-opted by insects to serve in the biochemical synthesis of a wide variety of compounds that function in communication. Chemical classification, although of interest, is of less value for many entomologists than the behaviors that the chemicals elicit. Very many behaviors of insects are governed by chemicals; nevertheless, we can distinguish pheromones that release specific behaviors from those that prime long-term, irreversible physiological changes. Thus, the stereotyped sexual behavior of a male moth is released by the female-emitted sex pheromone, whereas the crowding pheromone of locusts will prime maturation of gregarious phase individuals (section 6.10.5). Here, further classification of pheromones is based on five categories of behavior associated with sex, aggregation, spacing, trail forming, and alarm.
Sex pheromones
Male and female conspecific insects often communicate with chemical sex pheromones. Mate location and courtship may involve chemicals in two stages, with sex attraction pheromones acting at a distance, followed by close-up courtship pheromones employed prior to mating. The sex pheromones involved in attraction often differ from those used in courtship. Production and release of sex attractant pheromones tends to be restricted to the female, although there are lepidopterans and scorpionflies in which males are the releasers of distance attractants that lure females. The producer releases volatile pheromones that stimulate characteristic behavior in those members of the opposite sex within range of the odorous plume. An aroused recipient raises the antennae, orientates towards the source and walks or flies upwind to the source, often in a zig-zag track (Fig. 4.7) based on ability to respond rapidly to minor changes in pheromone concentration by direction change (Box 4.3). Each successive action appears to depend upon an increase in concentration of this airborne pheromone. As the insect approaches the source, cues such as sound and vision may be involved in close-up courtship behavior.
Courtship (section 5.2), which involves co-ordination of the two sexes, may require close-up chemical stimulation of the partner with a courtship pheromone. This pheromone may be simply a high concentration of the attractant pheromone, but “aphrodisiac” chemicals do exist, as seen in the queen butterfly (Nymphalidae: Danaus gilippus). The males of this species, as with several other lepidopterans, have extrusible abdominal hairpencils (brushes), which produce a pheromone that is dusted directly onto the antennae of the female, whilst both are in flight (Fig. 4.8). The effect of this pheromone is to placate a natural escape reaction of the female, who alights, folds her wings and allows copulation. In D. gilippus, this male courtship pheromone, a pyrrolixidine alkaloid called danaidone, is essential to successful courtship. However, the butterfly cannot synthesize it without acquiring the chemical precursor by feeding on selected plants as an adult. In the arctiid moth, Creatonotus gangis, the precursor of the male courtship pheromone likewise cannot be synthesized by the moth, but is sequestered by the larva in the form of a toxic alkaloid from the host plant. The larva uses the chemical in its defense and at metamorphosis the toxins are transferred to the adult. Both sexes use them as defensive compounds, with the male additionally converting them to his pheromone. This he emits from inflatable abdominal tubes, called coremata, whose development is regulated by the alkaloid pheromone precursor.
A spectacular example of deceitful sexual signaling occurs in bolas spiders, which do not build a web, but whirl a single thread terminating in a sticky globule towards their moth prey (like gauchos using a bolas to hobble cattle). The spiders lure male moths to within reach of the bolas using synthetic lures of sex-attractant pheromone cocktails. The proportions of the components vary according to the abundance of particular moth species available as prey. Similar principles are applied by humans to control pest insects using lures containing synthetic sex pheromones or other attractants (section 16.9). Certain chemical compounds (e.g. methyl eugenol), that either occur naturally in plants or can be synthesized in the laboratory, are used to lure male fruit flies (Tephritidae) for pest management purposes. These male lures are sometimes called parapheromones, probably because the compounds may be used by the flies as a component in the synthesis of their sex pheromones and have been shown to improve mating success, perhaps by enhancing the male’s sexual signals.
Sex pheromones once were thought to be unique, species-specific chemicals, but in reality often they are chemical blends. The same chemical (e.g. a particular 14-carbon chain alcohol) may be present in a range of related and unrelated species, but it occurs in a blend of different proportions with several other chemicals. An individual component may elicit only one part of the sex attraction behavior, or a partial or complete mixture may be required. Often the blend produces a greater response than any individual component, a synergism that is widespread in insects that produce pheromone mixtures. Chemical structural similarity of pheromones may indicate systematic relationship amongst the producers. However, obvious anomalies arise when identical or very similar pheromones are synthesized from chemicals derived from identical diets by unrelated insects.
Even if individual components are shared by many species, the mixture of pheromones is very often species-specific. It is evident that pheromones, and the stereotyped behaviors that they evoke, are highly significant in maintenance of reproductive isolation between species. The species-specificity of sex pheromones avoids cross-species mating before males and females come into contact.
Aggregation pheromones
The release of an aggregation pheromone causes conspecific insects of both sexes to crowd around the source of the pheromone. Aggregation may lead to increased likelihood of mating but, in contrast to many sex pheromones, both sexes may produce and respond to aggregation pheromones. The potential benefits provided by the response include security from predation, maximum utilization of a scarce food resource, overcoming of host resistance, or cohesion of social insects, as well as the chance to mate.
Aggregation pheromones are known in six insect orders, including cockroaches, but their presence and mode of action has been studied in most detail in Coleoptera, particularly in economically damaging species such as stored-grain beetles (from several families) and timber and bark beetles (Curculionidae: Scolytinae). A well-researched example of a complex suite of aggregation pheromones is provided by the Californian western pine beetle, Dendroctonus brevicomis (Scolytinae), which attacks ponderosa pine (Pinus ponderosa). On arrival at a new tree, colonizing females release the pheromone exo-brevicomin augmented by myrcene, a terpene originating from the damaged pine tree. Both sexes of western pine beetle are attracted by this mixture, and newly arrived males then add to the chemical mix by releasing another pheromone, frontalin. The cumulative lure of frontalin, exo-brevicomin, and myrcene is synergistic, i.e. greater than any one of these chemicals alone. The aggregation of many pine beetles overwhelms the tree’s defensive secretion of resins.
Spacing pheromones
There is a limit to the number of western pine beetles (D. brevicomis; see above) that attack a single tree. Cessation is assisted by reduction in the attractant aggregation pheromones, but deterrent chemicals also are produced. After the beetles mate on the tree, both sexes produce “anti-aggregation” pheromones called verbenone and trans-verbenone, and males also emit ipsdienol. These deter further beetles from landing close by, encouraging spacing out of new colonists. When the resource is saturated, further arrivals are repelled.
Such semiochemicals, called spacing, epideictic, or dispersion pheromones, may effect appropriate spacing on food resources, as with some phytophagous insects. Several species of tephritid flies lay eggs singly in fruit where the solitary larva is to develop. Spacing occurs because the ovipositing female deposits an oviposition-deterrent pheromone on the fruit on which she has laid an egg, thereby deterring subsequent oviposition. Social insects, which by definition are aggregated, utilize pheromones to regulate many aspects of their behavior, including the spacing between colonies. Spacer pheromones of colony-specific odors may be used to ensure an even dispersal of colonies of conspecifics, as in African weaver ants (Formicidae: Oecophylla longinoda).
Trail-marking pheromones
Many social insects use pheromones to mark their trails, particularly to food and the nest. Trail-marking pheromones are volatile and short-lived chemicals that evaporate within days unless reinforced (perhaps as a response to a food resource that is longer lasting than usual). Trail pheromones in ants are commonly metabolic waste products excreted by the poison gland. These need not be species-specific for several species share some common chemicals. Dufour’s gland secretions of some ant species may be more species-specific chemical mixtures associated with marking of territory and pioneering trails. Ant trails appear to be non-polar, i.e. the direction to nest or food resource cannot be determined by the trail odor.
In contrast to trails laid on the ground, an airborne trail — an odor plume — has directionality because of increasing concentration of the odor towards the source. An insect may rely upon angling the flight path relative to the direction of the wind that brings the odor, resulting in a zig-zag upwind flight towards the source. Each directional shift is produced where the odor diminishes at the edge of the plume (Fig. 4.7).
Alarm pheromones
Nearly two centuries ago it was recognized that workers of honey bees (Apis mellifera) were alarmed by a freshly extracted sting. In the intervening years many aggregating insects have been found to produce chemical releasers of alarm behavior — alarm pheromones — that characterize most social insects (termites and eusocial hymenopterans). In addition, alarm pheromones are known in several hemipterans, including subsocial treehoppers (Membracidae), aphids (Aphididae), and some other true bugs. Alarm pheromones are volatile, non-persistent compounds that are readily dispersed throughout the aggregation. Alarm is provoked by the presence of a predator, or in many social insects, a threat to the nest. The behavior elicited may be rapid dispersal, such as in hemipterans that drop from the host plant; or escape from an unwinnable conflict with a large predator, as in poorly defended ants living in small colonies. The alarm behavior of many eusocial insects is most familiar to us when disturbance of a nest induces many ants, bees, or wasps to an aggressive defense. Alarm pheromones attract aggressive workers and these recruits attack the cause of the disturbance by biting, stinging, or firing repellent chemicals. Emission of more alarm pheromone mobilizes further defenders. Alarm pheromone may be daubed over an intruder to aid in directing the attack.
Alarm pheromones may have been derived over evolutionary time from chemicals used as general antipredator devices (allomones; see below), utilizing glands co-opted from many different parts of the body to produce the substances. For example, hymenopterans commonly produce alarm pheromones from mandibular glands and also from poison glands, metapleural glands, the sting shaft, and even the anal area. All these glands also may be production sites for defensive chemicals.
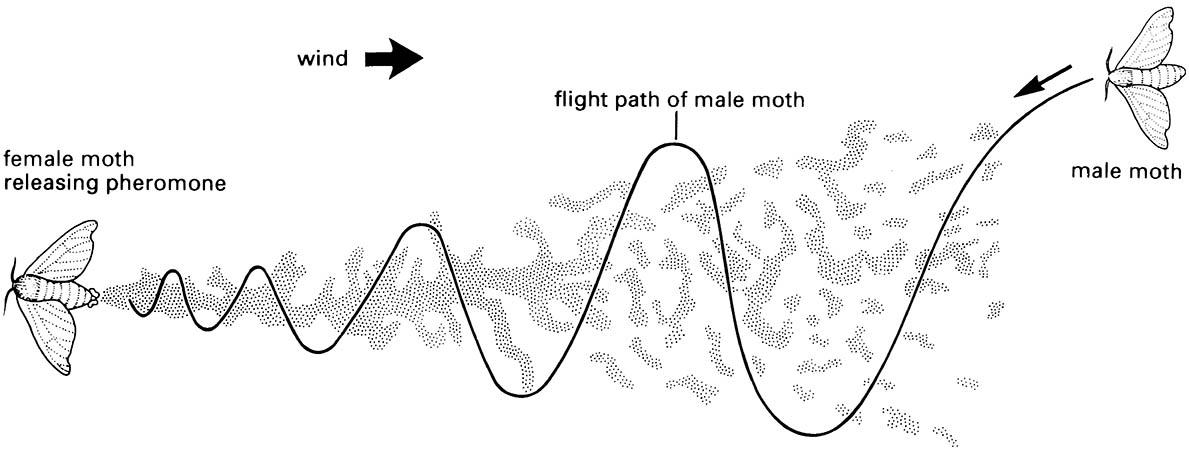
The pheromone trail forms a somewhat discontinuous plume because of turbulence, intermittent release, and other factors. (After Haynes & Birch 1985)
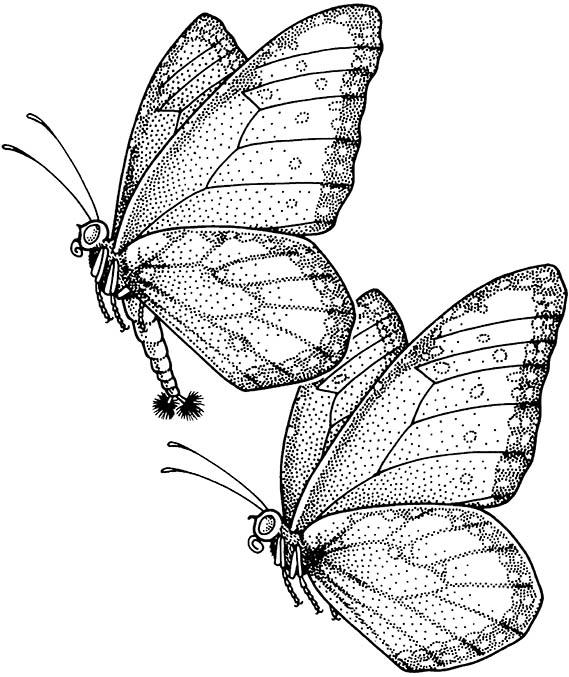
The male (above) has splayed hairpencils (at his abdominal apex) and is applying pheromone to the female (below). (After Brower et al. 1965)

