4.5. Insect behavior
Many of the insect behaviors mentioned in this chapter appear very complex, but behaviorists attempt to reduce them to simpler components. Thus, individual reflexes (simple responses to simple stimuli) can be identified, such as the flight response when the legs lose contact with the ground, and cessation of flight when contact is regained. Some extremely rapid reflex actions, such as the feeding lunge of odonate nymphs, or some “escape reactions” of many insects, depend upon a reflex involving giant axons that conduct impulses rapidly from sense organs to the muscles. The integration of multiple reflexes associated with movement of the insect may be divisible into:
- kinesis (plural: kineses), in which unorientated action varies according to stimulus intensity;
- taxis (plural: taxes), in which movement is directly towards or away from the stimulus.
Kineses include akinesis, unstimulated lack of movement, orthokinesis, in which speed depends upon stimulus intensity, and klinokinesis, which is a “random walk” with course changes (turns) being made when unfavorable stimuli are perceived and with the frequency of turns depending on the intensity of the stimulus. Increased exposure to unfavorable stimuli leads to increased tolerance (acclimation), so that random walking and acclimation will lead the insect to a favorable environment. The male response to the plume of sex attractant (Fig. 4.7) is an example of klinokinesis to a chemical stimulus. Ortho- and klinokineses are effective responses to diffuse stimuli, such as temperature or humidity, but different, more efficient responses are seen when an insect is confronted by less diffuse, gradient or point-source stimuli.
Kineses and taxes can be defined with respect to the type of stimulus eliciting a response. Appropriate prefixes include: anemo- for air currents, astro- for solar, lunar, or astral (including polarized light), chemo- for taste and odor, geo- for gravity, hygro- for moisture, phono- for sound, photo- for light, rheo- for water current, and thermo- for temperature. Orientation and movement may be positive or negative with respect to the stimulus source so that, for example, resistance to gravity is termed negative geotaxis, attraction to light is positive phototaxis, repulsion from moisture is negative hygrotaxis.
In klinotaxic behavior, an insect moves relative to a gradient (or cline) of stimulus intensity, such as a light source or a sound emission. The strength of the stimulus is compared on each side of the body by moving the receptors from side to side (as in head waving of ants when they follow an odor trail), or by detection of differences in stimulus intensity between the two sides of the body using paired receptors. The tympanal organs detect the direction of the sound source by differences in intensity between the two organs. Orientation with respect to a constant angle of light is termed menotaxis and includes the “light-compass” referred to in Box 4.4. Visual fixation of an object, such as prey, is termed telotaxis.
Often the relationship between stimulus and behavioral response is complex, as a threshold intensity may be required before an action ensues. A particular stimulatory intensity is termed a releaser for a particular behavior. Furthermore, complex behavior elicited by a single stimulus may comprise several sequential steps, each of which may have a higher threshold, requiring an increased stimulus. As described in section 4.3.2, a male moth responds to a low-level sex pheromone stimulus by raising the antennae; at higher levels he orientates towards the source; and at an even higher threshold, flight is initiated. Increasing concentration encourages continued flight and a second, higher threshold may be required before courtship ensues. In other behaviors, several different stimuli are involved, such as for courtship through to mating. This sequence can be seen as a long chain reaction of stimulus, action, new stimulus, next action, and so on, with each successive behavioral stage depending upon the occurrence of an appropriate new stimulus. An inappropriate stimulus during a chain reaction (such as the presentation of food while courting) is not likely to elicit the usual response.
Most insect behaviors are considered to be innate, i.e. they are programmed genetically to arise stereotypically upon first exposure to the appropriate stimulus. However, many behaviors are environmentally and physiologically modified: for example, virgins and mated females respond in very different ways to identical stimuli, and immature insects often respond to different stimuli compared with conspecific adults. Furthermore, experimental evidence shows that learning can modify innate behavior. By experimental teaching (using training and reward), bees and ants can learn to run a maze and butterflies can be induced to alter their favorite flower color. However, study of natural behavior (ethology) is more relevant to understanding the role played in the evolutionary success of insects’ behavioral plasticity, including the ability to modify behavior through learning. In pioneering ethological studies, Niko Tinbergen showed that a digger wasp (Crabronidae: Philanthus triangulum) can learn the location of its chosen nest site by making a short flight to memorize elements of the local terrain. Adjustment of prominent landscape features around the nest misleads the homing wasp. However, as wasps identify landmark relationships rather than individual features, the confusion may be only temporary. Closely related Bembix digger wasps (Sphecidae) learn nest locations through more distant and subtle markers, including the appearance of the horizon, and are not tricked by investigator ethologists moving local small-scale landmarks.
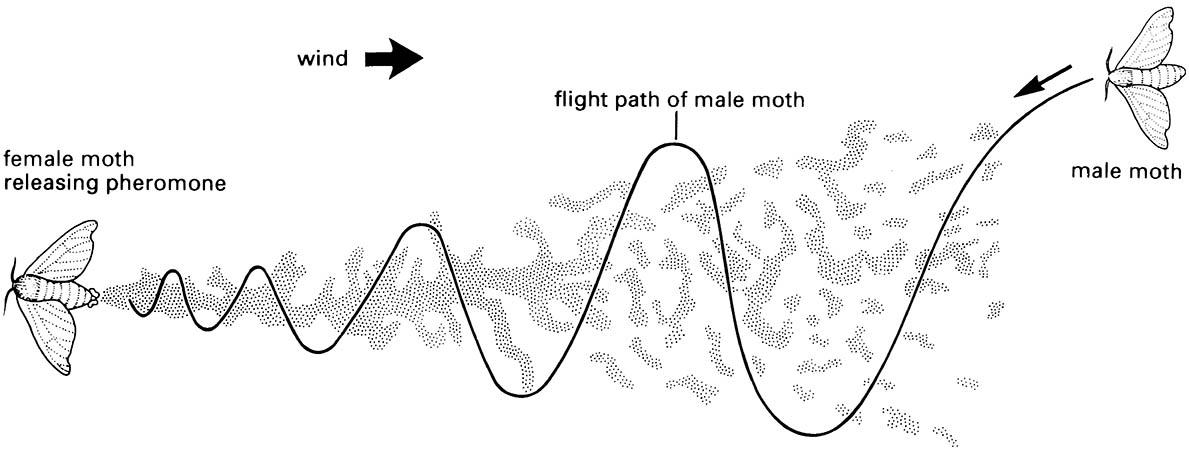
The pheromone trail forms a somewhat discontinuous plume because of turbulence, intermittent release, and other factors. (After Haynes & Birch 1985)

