4.1.4. Sound production
The commonest method of sound production by insects is by stridulation, in which one specialized body part, the scraper, is rubbed against another, the file. The file is a series of teeth, ridges, or pegs, which vibrate through contact with a ridged or plectrum-like scraper. The file itself makes little noise, and so has to be amplified to generate airborne sound. The horn-shaped bur- row of the mole cricket is an excellent sound enhancer (Fig. 4.5). Other insects produce many modifications of the body, particularly of wings and internal air sacs of the tracheal system, to produce amplification and resonance.
Sound production by stridulation occurs in some species of many orders of insects, but the Orthoptera show most elaboration and diversity. All stridulating orthopterans enhance their sounds using the tegmina (the modified fore wings). The file of katydids and crickets is formed from a basal vein of one or both tegmina, and rasps against a scraper on the other wing. Grasshoppers and locusts (Acrididae) rasp a file on the fore femora against a similar scraper on the tegmen.
Many insects lack the body size, power, or sophistication to produce high-frequency airborne sounds, but they can produce and transmit low-frequency sound by vibration of the substrate (such as wood, soil, or a host plant), which is a denser medium. Substrate vibrations are also a by-product of airborne sound production as in acoustic signaling insects, such as some katydids, whose whole body vibrates whilst producing audible airborne stridulatory sounds. Body vibrations, which are transferred through the legs to the substrate (plant or ground), are of low frequencies of 1–5000 Hz. Substrate vibrations can be detected by the female and appear to be used in closer range localization of the calling male, in contrast to the airborne signal used at greater distance.
A second means of sound production involves alternate muscular distortion and relaxation of a specialized area of elastic cuticle, the tymbal, to give individual clicks or variably modulated pulses of sound. Tymbal sound production is most audible to the human ear from cicadas, but many other hemipterans and some moths produce sounds from a tymbal. In the cicadas, only the males have these paired tymbals, which are located dorsolaterally, one on each side, on the first abdominal segment. The tymbal membrane is supported by a variable number of ribs. A strong tymbal muscle distorts the membrane and ribs to produce a sound; on relaxation, the elastic tymbal returns to rest. To produce sounds of high frequency, the tymbal muscle contracts asynchronously, with many contractions per nerve impulse (section 3.1.1). A group of chordonotal sensilla is present and a smaller tensor muscle controls the shape of the tymbal, thereby allowing alteration of the acoustic property. The noise of one or more clicks is emitted as the tymbal distorts, and further sounds may be produced during the elastic return on relaxation. The first abdominal segment contains air sacs — modified tracheae — tuned to resonate at or close to the natural frequency of tymbal vibration.
The calls of cicadas generally are in the range of 4–7 kHz, usually of high intensity, carrying as far as 1 km, even in thick forest. Sound is received by both sexes via tympanic membranes that lie ventral to the position of the male tymbal on the first abdominal segment. Cicada calls are species-specific — studies in New Zealand and North America show specificity of duration and cadence of introductory cueing phases inducing timed responses from a prospective mate. Interestingly however, song structures are very homoplasious, with similar songs found in distantly related taxa, but closely related taxa differing markedly in their song.
In other sound-producing hemipterans, both sexes may possess tymbals but because they lack abdominal air sacs, the sound is very damped compared with that of cicadas. The sounds produced by Nilaparvata lugens (the brown planthopper; Delphacidae), and probably other non-cicadan hemipterans, are transmitted by vibration of the substrate, and are specifically associated with mating.
Certain moths can hear the ultrasound produced by predatory bats, and moths themselves can produce sound using metepisternal tymbals. The high-frequency clicking sounds that arctiid moths produce can cause bats to veer away from attack, and may have the following (not mutually exclusive) roles:
- interspecific communication between moths;
- interference with bat sonar systems;
- aural mimicry of a bat to delude the predator about the presence of a prey item;
- warning of distastefulness (aposematism; see section 14.4).
The humming or buzzing sound characteristic of swarming mosquitoes, gnats, and midges is a flight tone produced by the frequency of wing beat. This tone, which can be virtually species-specific, differs between the sexes: the male produces a higher tone than the female. The tone also varies with age and ambient temperature for both sexes. Male insects that form nuptial (mating) swarms recognize the swarm site by species-specific environmental markers rather than audible cues (section 5.1); they are insensitive to the wing tone of males of their species. Neither can the male detect the wing tone of immature females — the Johnson’s organ in his antenna responds only to the wing tone of physiologically receptive females.
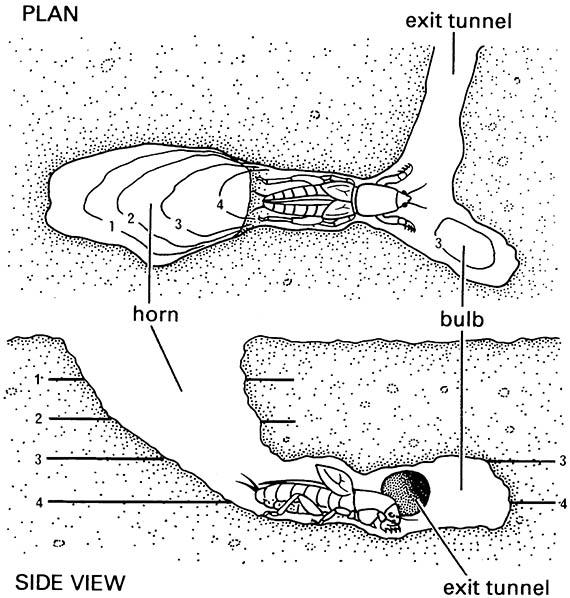
(After Bennet-Clark 1989)

