Box 4.3. Reception of communication molecules
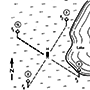
Pheromones, and indeed all signaling chemicals (semiochemicals), must be detectable in the smallest quantities. For example, the moth approaching a pheromone source portrayed in Fig. 4.7, must detect an initially weak signal, and then respond appropriately by orientating towards it, distinguishing abrupt changes in concentration ranging from zero to short-lived concentrated puffs. This involves a physiological ability to monitor continuously and respond to aerial pheromone levels in a process involving extra- and intracellular events.
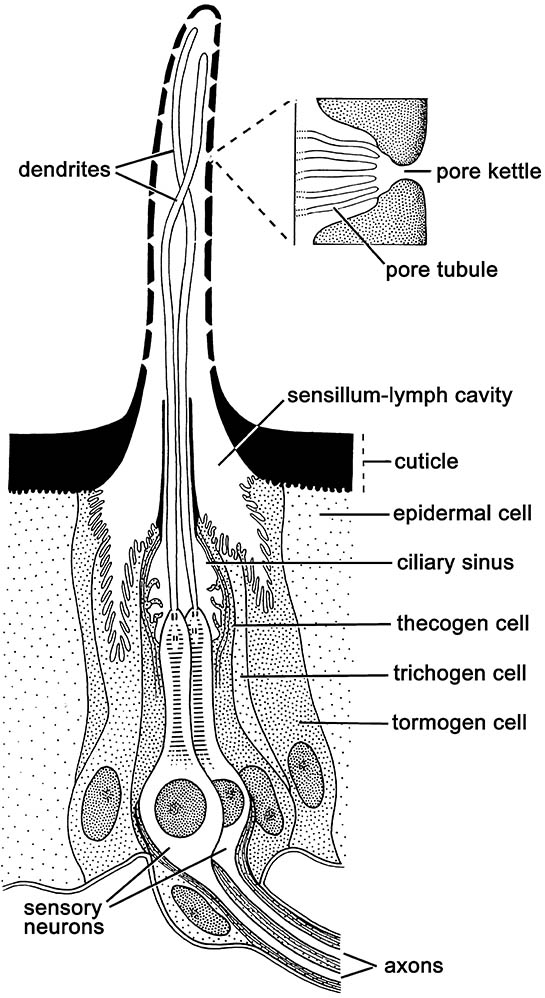
Ultrastructural studies of Drosophila melanogaster and several species of moth allow identification of several types of chemoreceptive (olfactory) sensilla: namely sensilla basiconica, sensilla trichodea, and sensilla coeloconica. These sensillar types are widely distributed across insect taxa and structures but most often are concentrated on the antenna. Each sensilla has from two to multiple subtypes which differ in their sensitivity and tuning to different communication chemicals. The structure of a generalized multiporous olfactory sensillum in the accompanying illustration follows Birch and Haynes (1982) and Zacharuk (1985).
To be detected, first the chemical must arrive at a pore of an olfactory sensillum. In a multiporous sensillum, it enters a pore kettle and contacts and crosses the cuticular lining of a pore tubule. Because pheromones (and other semiochemicals) largely are hydrophobic (lipophilic) compounds they must be made soluble to reach the receptors. This role falls to odorant-binding proteins (OBP) produced in the tormogen and trichogen cells (Fig. 4.1), from which they are secreted into the sensillum-lymph cavity that surrounds the dendrite of the receptor. Specific OBPs bind the semiochemical into a soluble ligand (OBP—pheromone complex) which is protected as it diffuses through the lymph to the dendrite surface. Here, interaction with negatively charged sites transforms the complex, releasing the pheromone to the binding site of the appropriate olfactory receptors located on the dendrite of the neuron, triggering a cascade of neural activity leading to appropriate behavior.
Much research has involved detection of pheromones because of their use in pest management (see section 16.9), but the principles revealed apparently apply to semiochemical reception across a range of organs and taxa. Thus, experiments with the electroantennogram (Box 4.2) using a single sensillum show highly specific responses to particular semiochemicals, and failure to respond even to “trivially” modified compounds. Studied OBPs appear to be one-to-one matched with each semiochemical, but insects apparently respond to more chemical cues than there are OBPs yet revealed. Additionally, olfactory receptors on the dendrite surface seemingly may be less specific, being triggered by a range of unrelated ligands. Furthermore, the model above does not address the frequently observed synergistic effects, in which a cocktail of chemicals provokes a stronger response than any component alone. It remains an open question as to exactly how insects are so spectacularly sensitive to so many specific chemicals, alone or in combination. This is an active research area, with microphysiology and molecular tools providing many new insights.
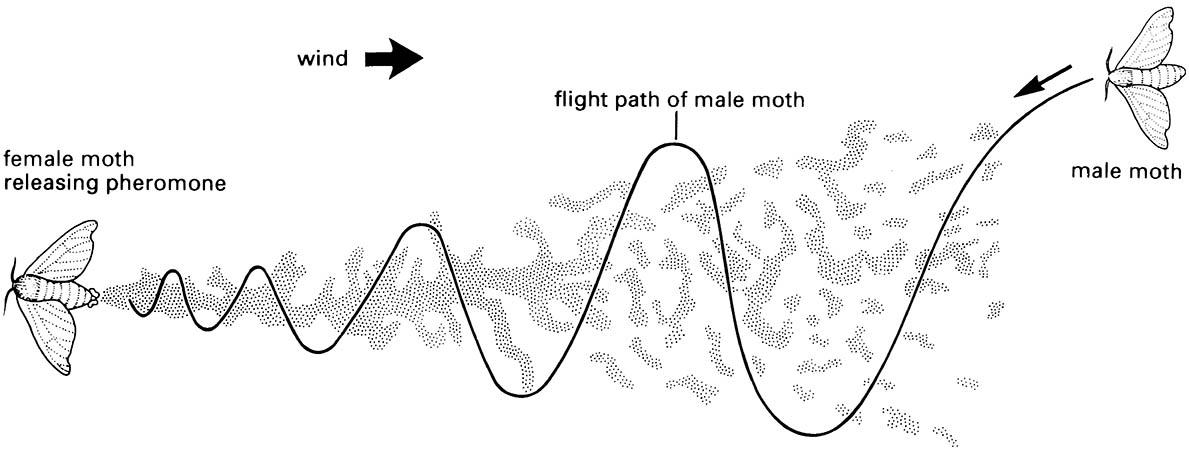
The pheromone trail forms a somewhat discontinuous plume because of turbulence, intermittent release, and other factors. (After Haynes & Birch 1985)
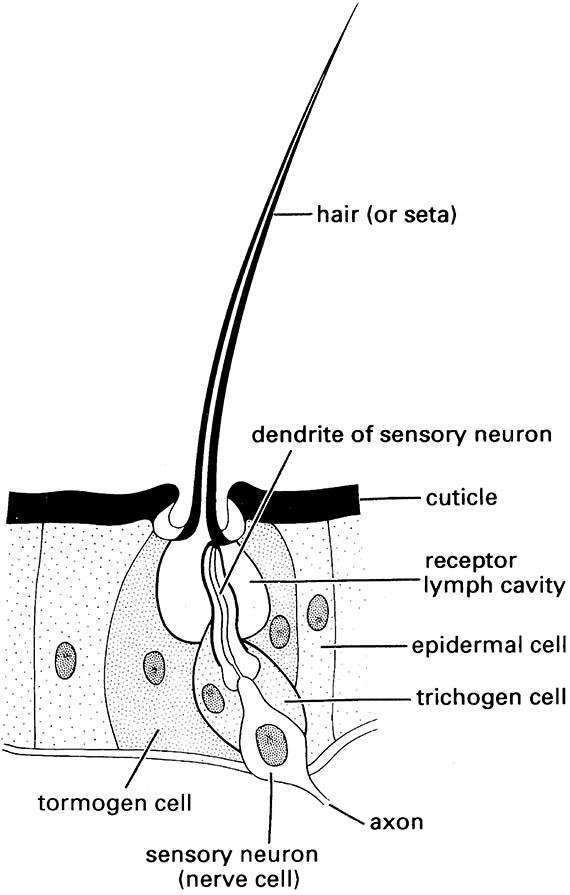
(After Chapman 1991)