4.4.4. Compound eyes
The most sophisticated insect visual organ is the compound eye. Virtually all adult insects and nymphs have a pair of large, prominent compound eyes, which often cover nearly 360 degrees of visual space.
The compound eye is based on repetition of many individual units called ommatidia (Fig. 4.10). Each ommatidium resembles a simple stemma: it has a cuticular lens overlying a crystalline cone, which directs and focuses light onto eight (or maybe 6–10) elongate retinula cells (see transverse section in Fig. 4.10). The retinula cells are clustered around the longitudinal axis of each ommatidium and each contributes a rhabdomere to the rhabdom at the center of the ommatidium. Each cluster of retinula cells is surrounded by a ring of light-absorbing pigment cells, which optically isolates an ommatidium from its neighbors.
The corneal lens and crystalline cone of each ommatidium focus light onto the distal tip of the rhabdom from a region about 2–5 degrees across. The field of view of each ommatidium differs from that of its neighbors and together the array of all ommatidia provides the insect with a panoramic image of the world. Thus, the actual image formed by the compound eye is of a series of apposed points of light of different intensities, hence the name apposition eye.
The light sensitivity of apposition eyes is limited severely by the small diameter of facet lenses. Crepuscular and nocturnal insects, such as moths and some beetles, overcome this limitation with a modified optical design of compound eyes, called optical superposition eyes. In these, ommatidia are not isolated optically from each other by pigment cells. Instead, the retina is separated by a wide clear zone from the corneal facet lenses, and many lenses co-operate to focus light on an individual rhabdom (light from many lenses super-imposes on the retina). The light sensitivity of these eyes is thus greatly enhanced. In some optical superposition eyes screening pigment moves into the clear zone during light adaptation and by this means the ommatidia become isolated optically as in the apposition eye. At low light levels, the screening pigment moves again towards the outer surface of the eye to open up the clear zone for optical superposition to occur.
Because the light arriving at a rhabdom has passed through many facet lenses, blurring is a problem in optical superposition eyes and resolution generally is not as good as in apposition eyes. However, high light sensitivity is much more important than good resolving power in crepuscular and nocturnal insects whose main concern is to see anything at all. In the eyes of some insects, photon-capture is increased even further by a mirror-like tapetum of small tracheae at the base of the retinula cells; this reflects light that has passed unabsorbed through a rhabdom, allowing it a second pass. Light reflecting from the tapetum produces the bright eye shine seen when an insect with an optical superposition eye is illuminated in the flashlight or car headlight beam at night.
In comparison with a vertebrate eye, the resolving power of insect compound eyes is rather unimpressive. However, for the purpose of flight control, navigation, prey capture, predator avoidance, and mate-finding they obviously do a splendid job. Bees can memorize quite sophisticated shapes and patterns, and flies and odonates hunt down prey insects or mates in extremely fast, aerobatic flight. Insects in general are exquisitely sensitive to image motion, which provides them with useful cues for avoiding obstacles and landing, and for distance judgment. Insects, however, cannot easily use binocular vision for the perception of distance because their eyes are so close together and their resolution is quite poor. A notable exception is the praying mantid, which is the only insect known to make use of binocular disparity to localize prey.
Within one ommatidium, most studied insects possess several classes of retinula cells that differ in their spectral sensitivities; this feature means that each responds best to light of a different wavelength. Variations in the molecular structure of visual pigments are responsible for these differences in spectral sensitivity and are a prerequisite for the color vision of flower visitors such as bees and butterflies. Some insects are pentachromats, with five classes of receptors of differing spectral sensitivities, compared with human di- or trichromats. Most insects can perceive ultraviolet light (which is invisible to us) allowing them to see distinctive alluring flower patterns visible only in the ultraviolet.
Light emanating from the sky and reflected light from water surfaces or shiny leaves is polarized, i.e. it has greater vibration in some planes than in others. Many insects can detect the plane of polarization of light and utilize this in navigation, as a compass or as an indicator of water surfaces. The pattern of polarized skylight changes its position as the sun moves across the sky, so that insects can use small patches of clear sky to infer the position of the sun, even when it is not visible. In like manner, an African dung beetle has been shown to orientate using polarized moonlight in the absence of direct sighting of the moon, perhaps representing a more general ability amongst nocturnal insects. The microvillar organization of the insect rhabdomere makes insect photoreceptors inherently sensitive to the plane of polarization of light, unless precautions are taken to scramble the alignment of microvilli. Insects with well-developed navigational abilities often have a specialized region of retina in the dorsal visual field, the dorsal rim, in which retinula cells are highly sensitive to the plane of polarization of light. Ocelli and stemmata also may be involved in the detection of polarized light.
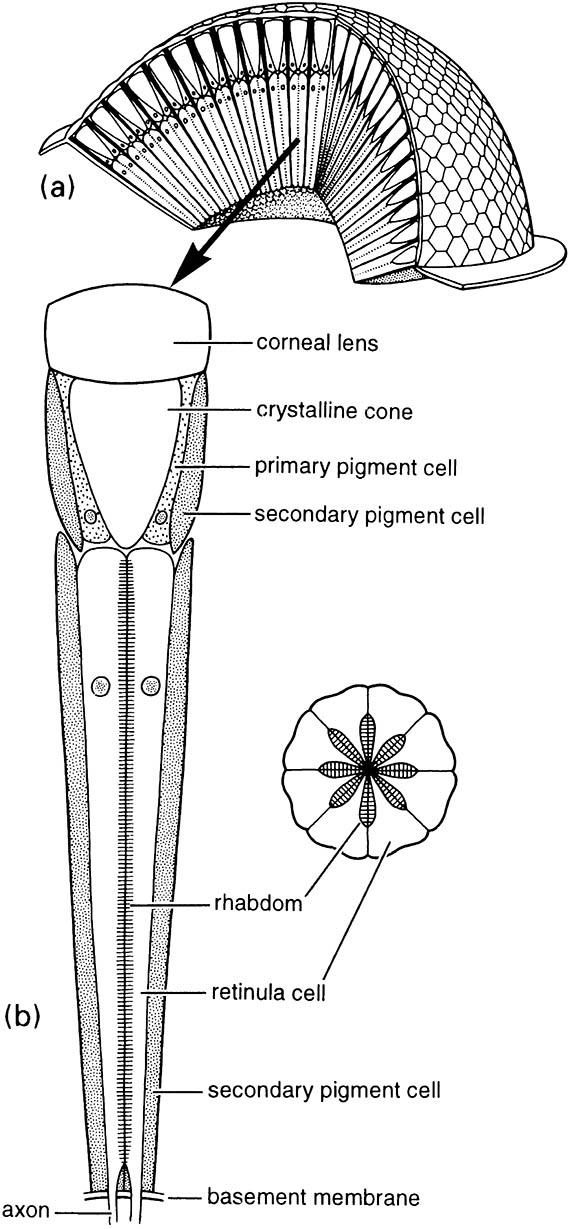
(a) a cutaway view showing the arrangement of the ommatidia and the facets; (b) a single ommatidium with an enlargement of a transverse section. (After CSIRO 1970; Rossel 1989)

