2.5.1. Terminalia
The anal-genital part of the abdomen, known as the terminalia, consists generally of segments 8 or 9 to the abdominal apex. Segments 8 and 9 bear the genitalia; segment 10 is visible as a complete segment in many “lower” insects but always lacks appendages; and the small segment 11 is represented by a dorsal epiproct and pair of ventral paraprocts derived from the sternum (Fig. 2.23b). A pair of appendages, the cerci, articulates laterally on segment 11; typically these are annulated and filamentous but have been modified (e.g. the forceps of earwigs) or reduced in different insect orders. An annulated caudal filament, the median appendix dorsalis, arises from the tip of the epiproct in apterygotes, most mayflies (Ephemeroptera), and a few fossil insects. A similar structure in nymphal stoneflies (Plecoptera) is of uncertain homology. These terminal abdominal segments have excretory and sensory functions in all insects, but in adults there is an additional reproductive function.
The organs concerned specifically with mating and the deposition of eggs are known collectively as the external genitalia, although they may be largely internal. The components of the external genitalia of insects are very diverse in form and often have considerable taxonomic value, particularly amongst species that appear structurally similar in other respects. The male external genitalia have been used widely to aid in distinguishing species, whereas the female external genitalia may be simpler and less varied. The diversity and species-specificity of genitalic structures are discussed in section 5.5.
The terminalia of adult female insects include internal structures for receiving the male copulatory organ and his spermatozoa (sections 5.4 and 5.6) and external structures used for oviposition (egg-laying; section 5.8). Most female insects have an egg-laying tube, or ovipositor; it is absent in Isoptera, Phthiraptera, many Plecoptera, and most Ephemeroptera. Ovipositors take two forms:
- true, or appendicular, formed from appendages of abdominal segments 8 and 9 (Fig. 2.23b);
- substitutional, composed of extensible posterior abdominal segments (Fig. 2.23a).
Substitutional ovipositors include a variable number of the terminal segments and clearly have been derived convergently several times, even within some orders. They occur in many insects, including most Lepidoptera, Coleoptera, and Diptera. In these insects, the terminalia are telescopic and can be extended as a slender tube, manipulated by muscles attached to apodemes of the modified terga (Fig. 2.23a) and/or sterna.
Appendicular ovipositors represent the primitive condition for female insects and are present in Archaeognatha, Zygentoma, many Odonata, Orthoptera, some Hemiptera, some Thysanoptera, and Hymenoptera. In some Hymenoptera, the ovipositor is modified as a poison-injecting sting (Fig. 14.11) and the eggs are ejected at the base of the sting. In all other cases, the eggs pass down a canal in the shaft of the ovipositor (section 5.8). The shaft is composed of three pairs of valves (Fig. 2.23b,c) supported on two pairs of valvifers — the coxae + trochanters, or gonocoxites, of segments 8 and 9 (Fig. 2.23b). The gonocoxites of segment 8 have a pair of trochanteral endites (inner lobe from each trochanter), or gonapophyses, which form the first valves, whereas the gonocoxites of segment 9 have a pair of gonapophyses (the second valves) plus a pair of gonostyles (the third valves) derived from the distal part of the appendages of segment 9 (and homologous with the styles of the apterygotes mentioned above). In each half of the ovipositor, the second valve slides in a tongue-and-groove fashion against the first valve (Fig. 2.23c), whereas the third valve generally forms a sheath for the other valves.
The external genitalia of male insects include an organ for transferring the spermatozoa (either packaged in a spermatophore, or free in fluid) to the female and often involve structures that grasp and hold the partner during mating. Numerous terms are applied to the various components in different insect groups and homologies are difficult to establish. Males of Archaeognatha, Zygentoma, and Ephemeroptera have relatively simple genitalia consisting of gonocoxites, gonostyles, and sometimes gonapophyses on segment 9 (and also on segment 8 in Archaeognatha), as in the female, except with a median penis (phallus) or, if paired or bilobed, penes, on segment 9 (Fig. 2.24a). The penis (or penes) is believed to be derived from the fused inner lobes (endites) of either the ancestral coxae or trochanters of segment 9. In the orthopteroid orders, the gonocoxites are reduced or absent, although gonostyles may be present (called styles), and there is a median penis with a lobe called a phallomere on each side of it. The evolutionary fate of the gonapophyses and the origin of the phallomeres are uncertain. In the “higher” insects — the hemipteroids and the holometabolous orders — the homologies and terminology of the male structures are even more confusing if one tries to compare the terminalia of different orders. The whole copulatory organ of higher insects generally is known as the aedeagus (edeagus) and, in addition to insemination, it may clasp and provide sensory stimulation to the female. Typically, there is a median tubular penis (although sometimes the term “aedeagus” is restricted to this lobe), which often has an inner tube, the endophallus, that is everted during insemination (Fig. 5.4b). The ejaculatory duct opens at the gonopore, either at the tip of the penis or the endophallus. Lateral to the penis is a pair of lobes or parameres, which may have a clasping and/or sensory function. Their origin is uncertain; they may be homologous with the gonocoxites and gonostyles of lower insects, with the phallomeres of orthopteroid insects, or be derived de novo, perhaps even from segment 10. This trilobed type of aedeagus is well exemplified in many beetles (Fig. 2.24b), but modifications are too numerous to describe here.
Much variation in male external genitalia correlates with mating position, which is very variable between and sometimes within orders. Mating positions include end-to-end, side-by-side, male below with his dorsum up, male on top with female dorsum up, and even venter-to-venter. In some insects, torsion of the terminal segments may take place post-metamorphosis or just prior to or during copulation, and asymmetries of male clasping structures occur in many insects. Copulation and associated behaviors are discussed in more detail in Chapter 5.
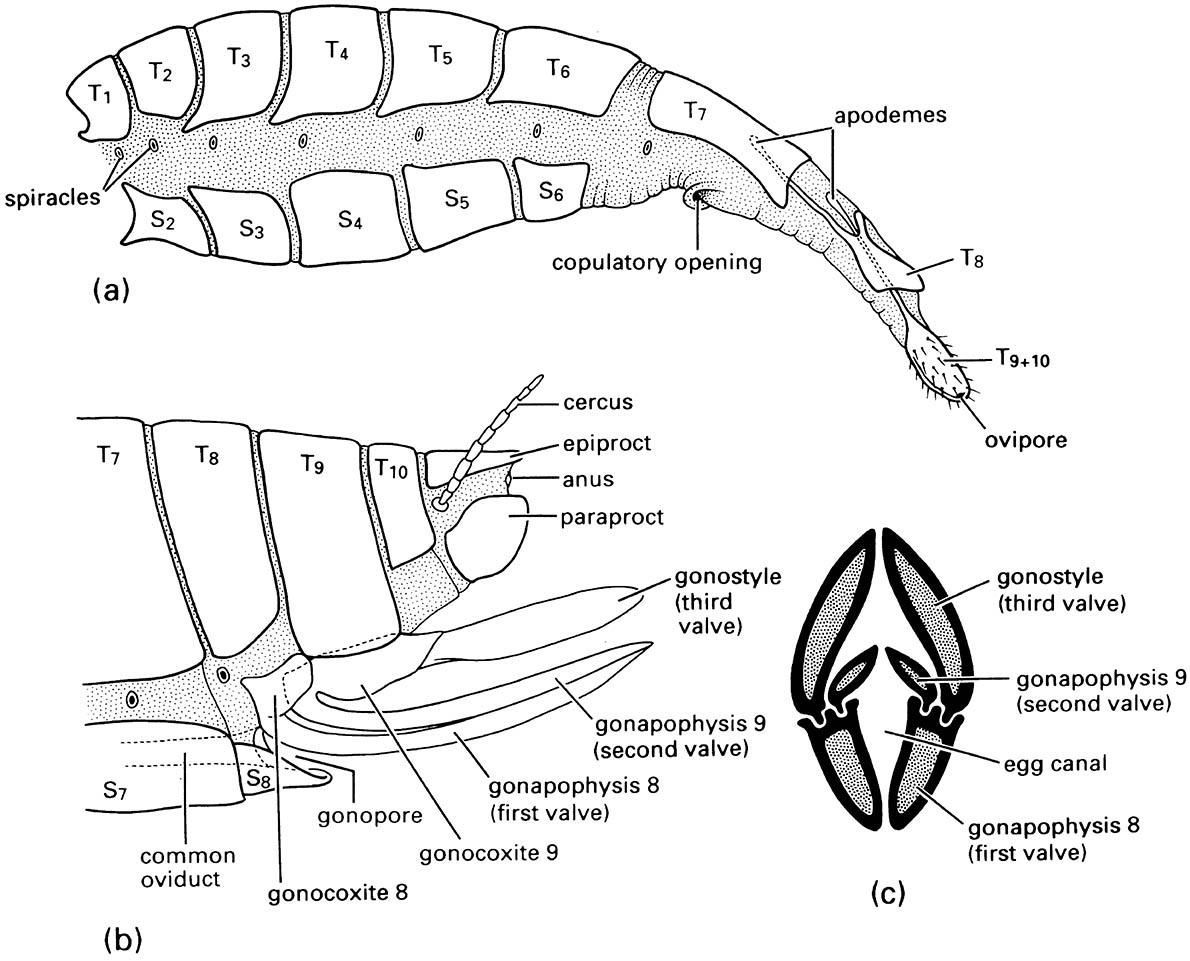
(a) lateral view of the abdomen of an adult tussock moth (Lepidoptera: Lymantriidae) showing the substitutional ovipositor formed from the extensible terminal segments; (b) lateral view of a generalized orthopteroid ovipositor composed of appendages of segments 8 and 9; (c) transverse section through the ovipositor of a katydid (Orthoptera: Tettigoniidae). T1—T10, terga of first to tenth segments; S2—S8, sterna of second to eighth segments. ((a) After Eidmann 1929; (b) after Snodgrass 1935; (c) after Richards & Davies 1959)
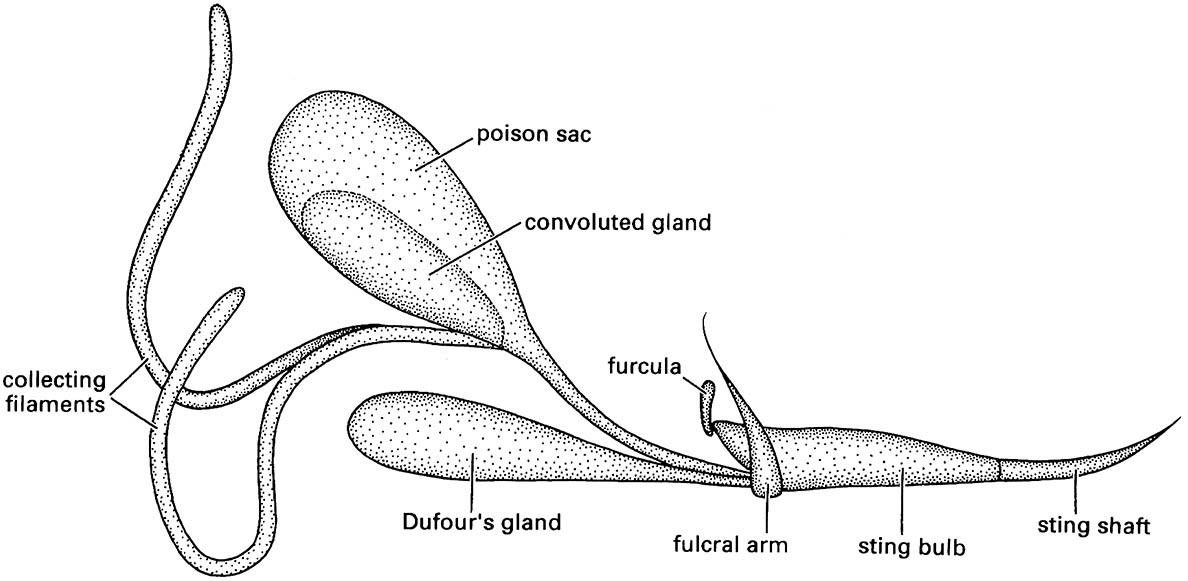
(After Hermann & Blum 1981)
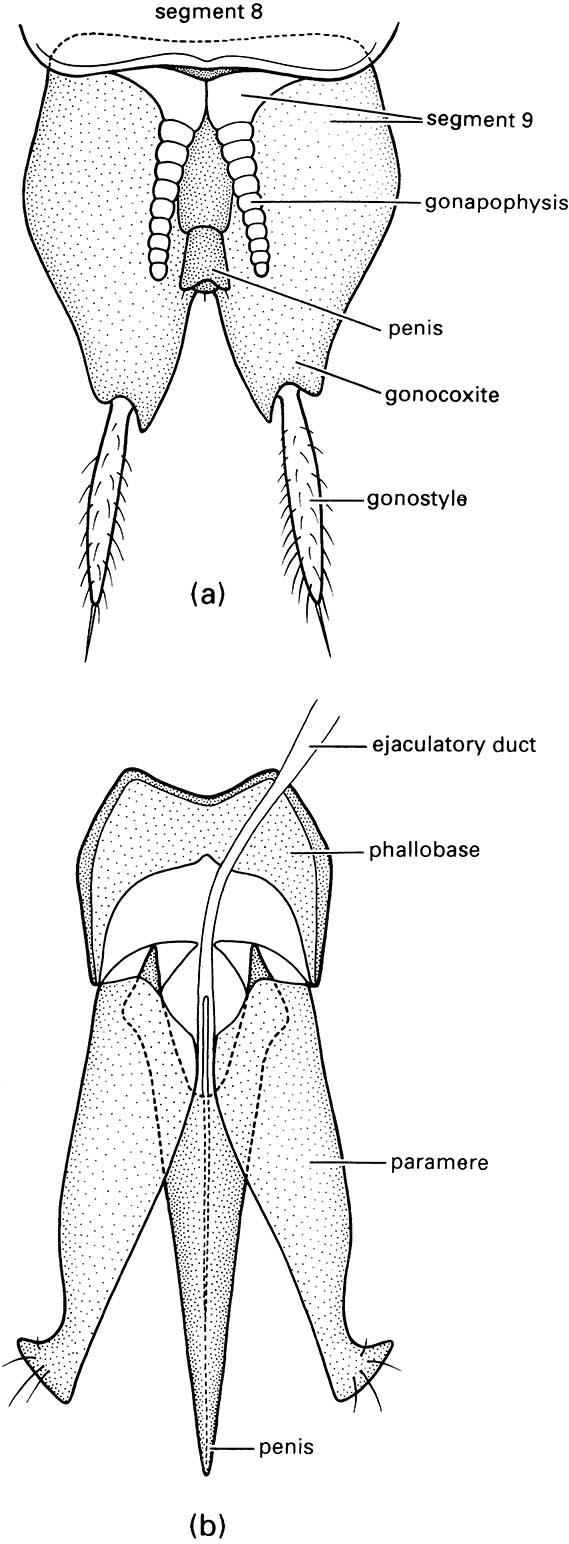
(a) Abdominal segment 9 of the bristletail Machilis variabilis (Archaeognatha: Machilidae). (b) Aedeagus of a click beetle (Coleoptera: Elateridae). ((a) After Snodgrass 1957)
Mating commences with the pair facing in the same direction, then the male rotates his eighth abdominal segment (90°) and genital capsule (180°), erects the aedeagus and gains entry to the female’s genital chamber, before he swings around to face in the opposite direction. The bugs may copulate for several hours, during which they walk around with the female leading and the male walking backwards. (a) Lateral view of the terminal segments, showing the valves of the female’s ovipositor in the male genital chamber; (b) longitudinal section showing internal structures of the reproductive system, with the tip of the male’s aedeagus in the fem ale’s spermatheca. (After Bonhag & Wick 1953)

