2.4.1. Legs
In most adult and nymphal insects, segmented fore, mid, and hind legs occur on the prothorax, mesothorax, and metathorax, respectively. Typically, each leg has six segments (Fig. 2.19) and these are, from proximal to distal: coxa, trochanter, femur, tibia, tarsus, and pretarsus (or more correctly posttarsus) with claws. Additional segments — the prefemur, patella, and basitarsus (Fig. 8.4a) — are recognized in some fossil insects and other arthropods, such as arachnids, and one or more of these segments are evident in some Ephemeroptera and Odonata. Primitively, two further segments lie proximal to the coxa and in extant insects one of these, the epicoxa, is associated with the wing articulation, or tergum, and the other, the subcoxa, with the pleuron (Fig. 8.4a).
The tarsus is subdivided into five or fewer components, giving the impression of segmentation; but, because there is only one tarsal muscle, tarsomere is a more appropriate term for each “pseudosegment”. The first tarsomere sometimes is called the basitarsus, but should not be confused with the segment called the basitarsus in certain fossil insects. The underside of the tarsomeres may have ventral pads, pulvilli, also called euplantulae, which assist in adhesion to surfaces. Terminally on the leg, the small pretarsus (enlargement in Fig. 2.19) bears a pair of lateral claws (also called ungues) and usually a median lobe, the arolium. In Diptera there may be a central spine-like or pad-like empodium (plural: empodia) which is not the same as the arolium, and a pair of lateral pulvilli (as shown for the bush fly, Musca vetustissima, depicted on the right side of the vignette of this chapter). These structures allow flies to walk on walls and ceilings. The pretarsus of Hemiptera may bear a variety of structures, some of which appear to be pulvilli, whereas others have been called empodia or arolia, but the homologies are uncertain. In some beetles, such as Coccinellidae, Chrysomelidae, and Curculionidae, the ventral surface of some tarsomeres is clothed with adhesive setae that facilitate climbing. The left side of the vignette for this chapter shows the underside of the tarsus of the leaf beetle Rhyparida (Chrysomelidae).
Generally the femur and tibia are the longest leg segments but variations in the lengths and robustness of each segment relate to their functions. For example, walking (gressorial) and running (cursorial) insects usually have well-developed femora and tibiae on all legs, whereas jumping (saltatorial) insects such as grasshoppers have disproportionately developed hind femora and tibiae. In aquatic beetles (Coleoptera) and bugs (Hemiptera), the tibiae and/or tarsi of one or more pairs of legs usually are modified for swimming (natatorial) with fringes of long, slender hairs. Many ground-dwelling insects, such as mole crickets (Orthoptera: Gryllotalpidae), nymphal cicadas (Hemiptera: Cicadidae), and scarab beetles (Scarabaeidae), have the tibiae of the fore legs enlarged and modified for digging (fossorial) (Fig. 9.2), whereas the fore legs of some predatory insects, such as mantispid lacewings (Neuroptera) and mantids (Mantodea), are specialized for seizing prey (raptorial) (Fig. 13.3). The tibia and basal tarsomere of each hind leg of honey bees are modified for the collection and carriage of pollen (Fig. 12.4).
These “typical” thoracic legs are a distinctive feature of insects, whereas abdominal legs are confined to the immature stages of holometabolous insects. There have been conflicting views on whether (i) the legs on the immature thorax of the Holometabola are develop- mentally identical (serially homologous) to those of the abdomen, and/or (ii) the thoracic legs of the holometabolous immature stages are homologous with those of the adult. Detailed study of musculature and innervation shows similarity of development of thoracic legs throughout all stages of insects with ametaboly (without metamorphosis, as in silverfish) and hemimetaboly (partial metamorphosis and no pupal stage) and in adult Holometabola, having identical innervation through the lateral nerves. Moreover, the oldest known larva (from the Upper Carboniferous) has thoracic and abdominal legs/leglets each with a pair of claws, as in the legs of nymphs and adults. Although larval legs appear similar to those of adults and nymphs, the term prolegs is used for the larval leg. Prolegs on the abdomen, especially on caterpillars, usually are lobelike and each bears an apical circle or band of small sclerotized hooks, or crochets. The thoracic prolegs may possess the same number of segments as the adult leg, but the number is more often reduced, apparently through fusion. In other cases, the thoracic prolegs, like those of the abdomen, are unsegmented outgrowths of the body wall, often bearing apical hooks.
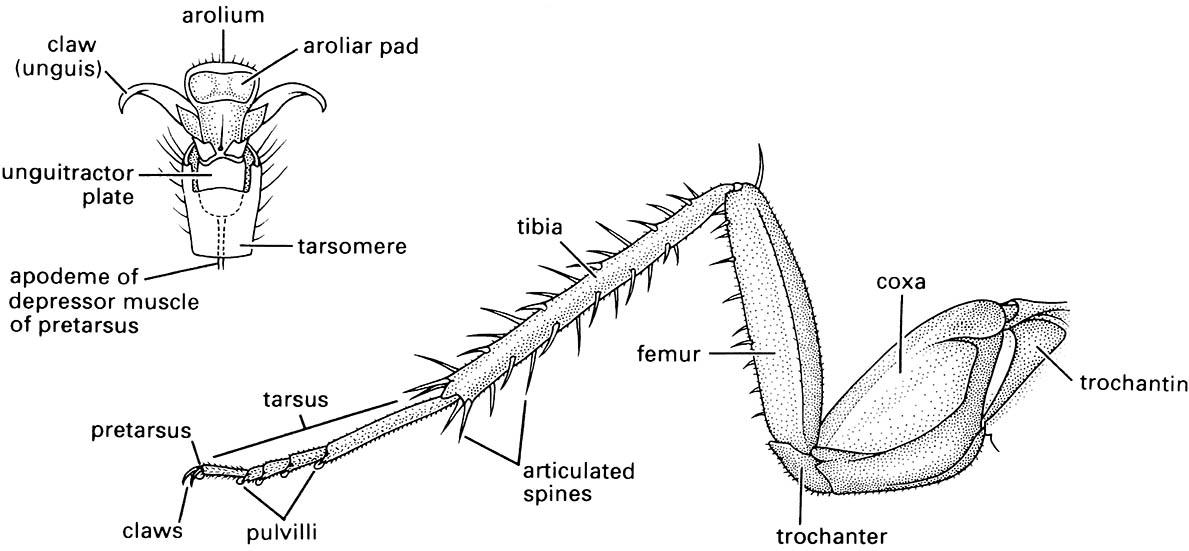
(After Cornwell 1968; enlargement after Snodgrass 1935)
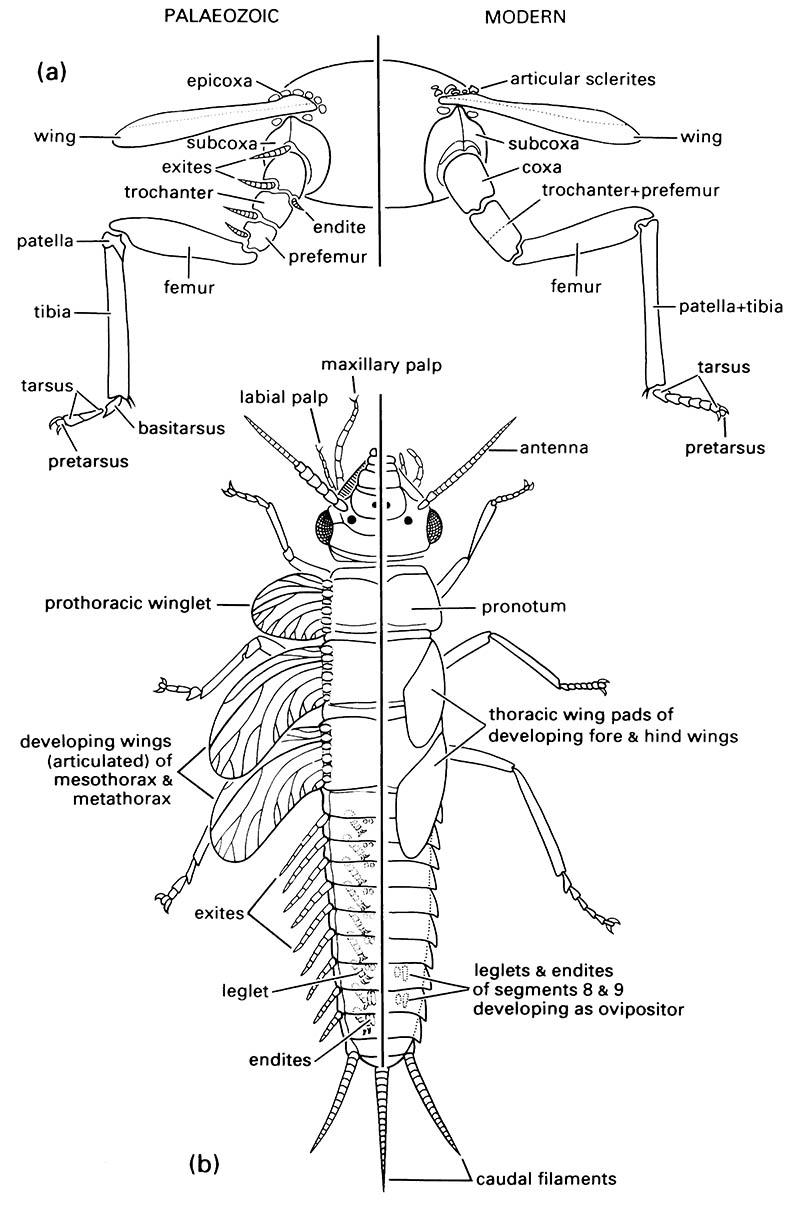
(a) thoracic segment of adult showing generalized condition of appendages; (b) dorsal view of nymphal morphology. (Modified from Kukalová-Peck 1991; to incorporate ideas of J.W.H. Trueman (unpublished))
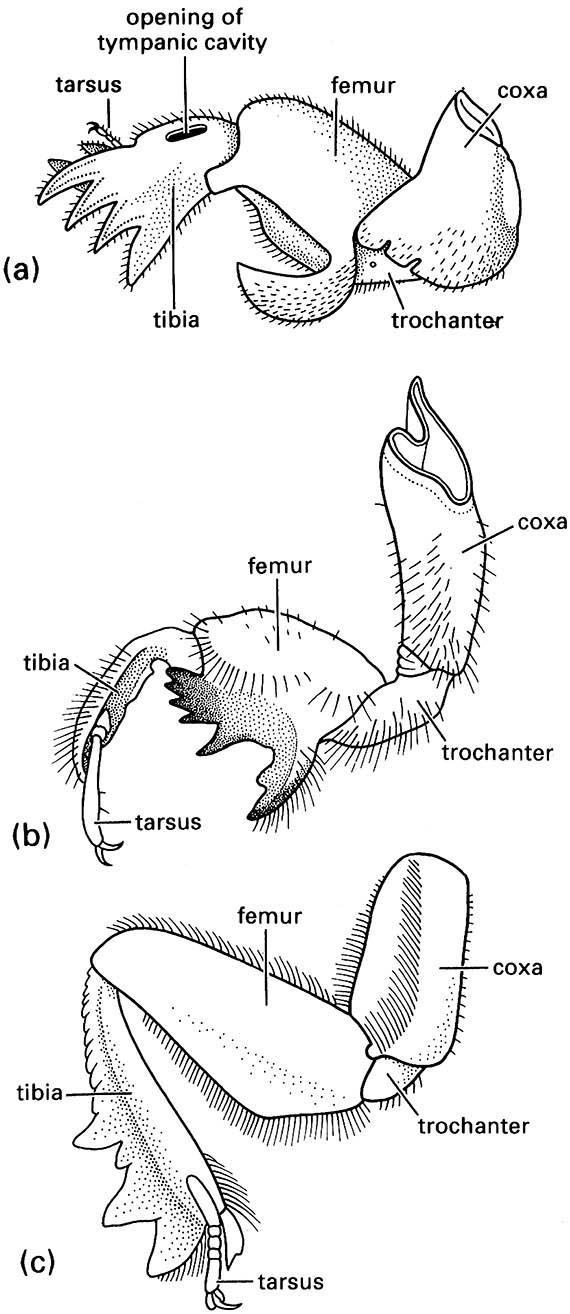
(a) a mole cricket of Gryllotalpa (Orthoptera: Gryllotalpidae); (b) a nymphal periodical cicada of Magicicada (Hemiptera: Cicadidae); and (c) a scarab beetle of Canthon (Coleoptera: Scarabaeidae). ((a) After Frost 1959; (b) after Snodgrass 1967; (c) after Richards & Davies 1977)
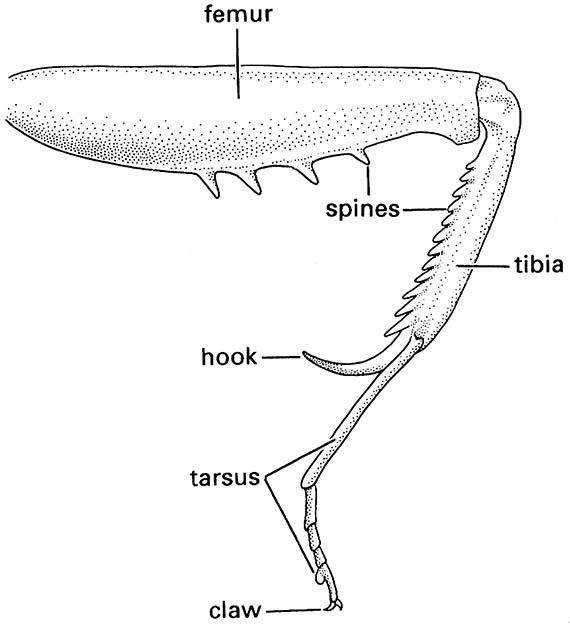
(After Preston-Mafham 1990)
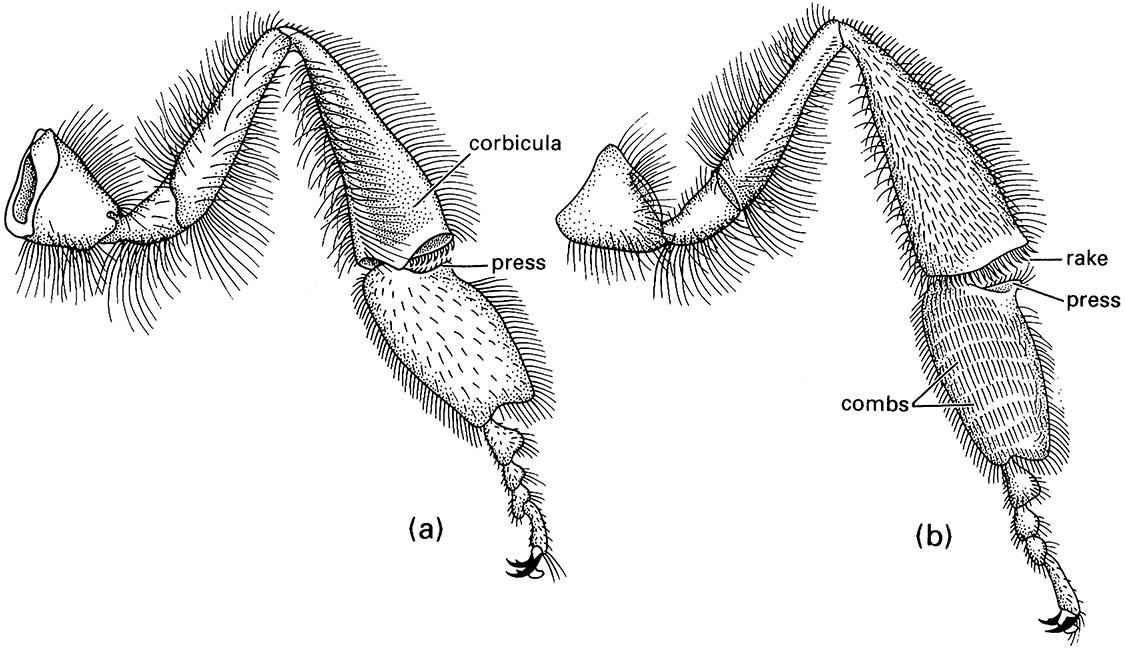
(a) outer surface showing corbicula, or pollen basket (consisting of a depression fringed by stiff setae), and the press that pushes the pollen into the basket; (b) the inner surface with the combs and rakes that manipulate pollen into the press prior to packing. (After Snodgrass 1956; Winston 1987)

