Box 4.4. Biological clocks
Seasonal changes in environmental conditions allow insects to adjust their life histories to optimize the use of suitable conditions and minimize the impact of unsuitable ones (e.g. through diapause; section 6.5). Similar physical fluctuations on a daily scale encourage a diurnal (daily) cycle of activity and quiescence. Nocturnal insects are active at night, diurnal ones by day, and crepuscular insect activity occurs at dusk and dawn when light intensities are transitional. The external physical environment, such as light—dark or temperature, controls some daily activity patterns, called exogenous rhythms. However, many other periodic activities are internally driven endogenous rhythms that have a clock-like or calendar-like frequency irrespective of external conditions. Endogenous periodicity is frequently about 24 h (circadian), but lunar and tidal periodicities govern the emergence of adult aquatic midges from large lakes and the marine intertidal zones, respectively. This unlearned, once-in-a-lifetime rhythm which allows synchronization of eclosion demonstrates the innate ability of insects to measure passing time.
Experimentation is required to discriminate between exogenous and endogenous rhythms. This involves observing what happens to rhythmic behavior when external environmental cues are altered, removed, or made invariate. Such experiments show that inception (setting) of endogenous rhythms is found to be day length, with the clock then free-running, without daily reinforcement by the light—dark cycle, often for a considerable period. Thus, if nocturnal cockroaches that become active at dusk are kept at constant temperature in constant light or dark, they will maintain the dusk commencement of their activities at a circadian rhythm of 23–25 h. Rhythmic activities of other insects may require an occasional clock-setting (such as darkness) to prevent the circadian rhythm drifting, either through adaptation to an exogenous rhythm or into arrhythmy.
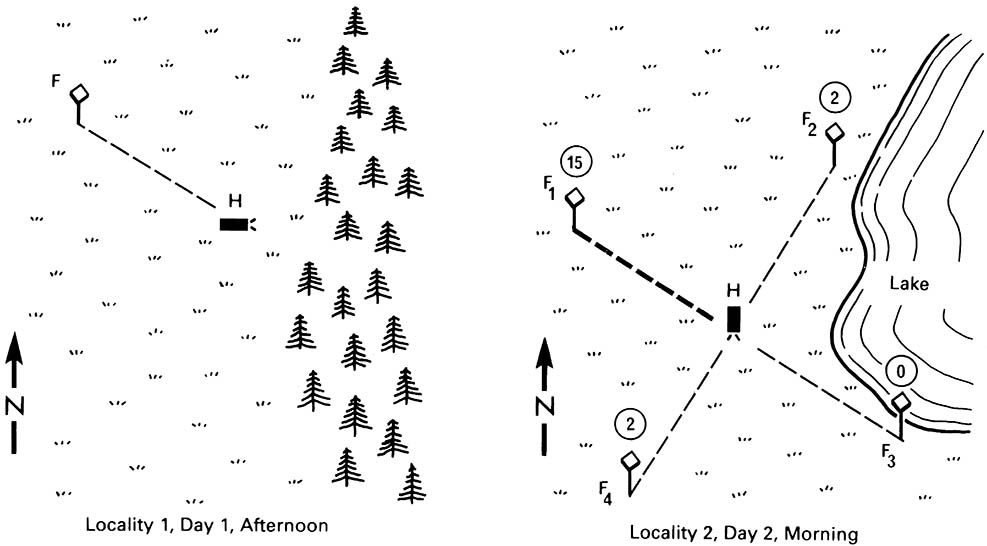
Biological clocks allow solar orientation — the use of the sun’s elevation above the horizon as a compass — provided that there is a means of assessing (and compensating for) the passage of time. Some ants and honey bees use a “light-compass”, finding direction from the sun’s elevation and using the biological clock to compensate for the sun’s movement across the sky. Evidence came from an elegant experiment with honey bees trained to forage in the late afternoon at a feeding table (F) placed 180 m NW of their hive (H), as depicted in the left figure (after Lindauer 1960). Overnight the hive was moved to a new location to prevent use of familiar landmarks in foraging, and a selection of four feeding tables (F 1-4) was provided at 180 m, NW, SW, SE, and NE from the hive. In the morning, despite the sun being at a very different angle to that during the afternoon training, 15 of the 19 bees were able to locate the NW table (as depicted in the figure on the right). The honey bee “dance language” that communicates direction and distance of food to other workers (Box 12.1) depends upon the capacity to calculate direction from the sun.
The circadian pacemaker (oscillator) that controls the rhythm is located in the brain; it is not an external photoperiod receptor. Experimental evidence shows that in cockroaches, beetles, and crickets a pacemaker lies in the optic lobes, whereas in some silkworms it lies in the cerebral lobes of the brain. In the well-studied Drosophila , a major oscillator site appears to be located between the lateral protocerebellum and the medulla of the optic. However, visualization of the sites of period gene activity is not localized, and there is increasing evidence of multiple pacemaker centers located through- out the tissues. Whether they communicate with each other or run independently is not yet clear.