11.2. Phytophagy (or herbivory)
The majority of plant species support complex faunas of herbivores, each of which may be defined in relation to the range of plant taxa used. Thus, monophages are specialists that feed on one plant taxon, oligophages feed on few, and polyphages are generalists that feed on many plant groups. The adjectives for these feeding categories are monophagous, oligophagous, and polyphagous. Gall-inducing cynipid wasps (Hymenoptera) exemplify monophagous insects as nearly all species are host-plant specific; furthermore, all cynipid wasps of the tribe Rhoditini induce their galls only on roses (Rosa) (Fig. 11.5d) and almost all species of Cynipini form their galls only on oaks (Quercus) (Fig. 11.5c). The monarch or wanderer butterfly, Danaus plexippus (Nymphalidae), is an example of an oligophagous insect, with larvae that feed on various milkweeds, predominantly species of Asclepias. The polyphagous gypsy moth, Lymantria dispar (Lymantriidae), feeds on a wide range of tree genera and species, and the Chinese wax scale, Ceroplastes sinensis (Hemiptera: Coccidae), is truly polyphagous with its recorded host plants belonging to about 200 species in at least 50 families. In general, most phytophagous insect groups, except Orthoptera, tend to be specialized in their feeding.
Many plants appear to have broad-spectrum defenses against a very large suite of enemies, including insect and vertebrate herbivores and pathogens. These primarily physical or chemical defenses are discussed in section 16.6 in relation to host-plant resistance to insect pests. Spines or pubescence on stems and leaves, silica or sclerenchyma in leaf tissue, or leaf shapes that aid camouflage are amongst the physical attributes of plants that may deter some herbivores. Furthermore, in addition to the chemicals considered essential to plant function, most plants contain compounds whose role generally is assumed to be defensive, although these chemicals may have, or once may have had, other metabolic functions or simply be metabolic waste products. Such chemicals are often called secondary plant compounds, noxious phytochemicals, or allelochemicals. A huge array exists, including phenolics (such as tannins), terpenoid compounds (essential oils), alkaloids, cyanogenic glycosides, and sulfur-containing glucosinolates. The anti-herbivore action of many of these compounds has been demonstrated or inferred. For example, in Acacia, the loss of the otherwise widely distributed cyanogenic glycosides in those species that harbor mutualistic stinging ants implies that the secondary plant chemicals do have an antiherbivore function in those many species that lack ant defenses.
In terms of plant defense, secondary plant compounds may act in one of two ways. At a behavioral level, these chemicals may repel an insect or inhibit feeding and/or oviposition. At a physiological level, they may poison an insect or reduce the nutritional content of its food. However, the same chemicals that repel some insect species may attract others, either for oviposition or feeding (thus acting as kairomones; section 4.3.3). Such insects, thus attracted, are said to be adapted to the chemicals of their host plants, either by tolerating, detoxifying, or even sequestering them. An example is the monarch butterfly, D. plexippus, which usually oviposits on milkweed plants, many of which contain toxic cardiac glycosides (cardenolides), which the feeding larva can sequester for use as an anti-predator device (sections 14.4.3 & 14.5.1).
Secondary plant compounds have been classified into two broad groups based on their inferred biochemical actions: (i) qualitative or toxic, and (ii) quantitative. The former are effective poisons in small quantities (e.g. alkaloids, cyanogenic glycosides), whereas the latter are believed to act in proportion to their concentration, being more effective in greater amounts (e.g. tannins, resins, silica). In practice, there probably is a continuum of biochemical actions, and tannins are not simply digestion-reducing chemicals but have more complex anti-digestive and other physiological effects. However, for insects that are specialized to feed on particular plants containing any secondary plant compound(s), these chemicals actually can act as phagostimulants. Furthermore, the narrower the host-plant range of an insect, the more likely that it will be repelled or deterred by non-host-plant chemicals, even if these substances are not noxious if ingested.
The observation that some kinds of plants are more susceptible to insect attack than others also has been explained by the relative apparency of the plants. Thus, large, long-lived, clumped trees are very much more apparent to an insect than small, annual, scattered herbs. Apparent plants tend to have quantitative secondary compounds, with high metabolic costs in their production. Unapparent plants often have qualitative or toxic secondary compounds, produced at little metabolic cost. Human agriculture often turns unapparent plants into apparent ones, when monocultures of annual plants are cultivated, with corresponding increases in insect damage.
Another consideration is the predictability of resources sought by insects, such as the suggested predictability of the presence of new leaves on a eucalypt tree or creosote bush in contrast to the erratic spring flush of new leaves on a deciduous tree. However, the question of what is predictability (or apparency) of plants to insects is essentially untestable. Furthermore, insects can optimize the use of intermittently abundant resources by synchronizing their life cycles to environmental cues identical to those used by the plant.
A third correlate of variation in herbivory rates concerns the nature and quantities of resources (i.e. light, water, nutrients) available to plants. One hypothesis is that insect herbivores feed preferentially on stressed plants (e.g. affected by water-logging, drought, or nutrient deficiency), because stress can alter plant physiology in ways beneficial to insects. Alternatively, insect herbivores may prefer to feed on vigorously growing plants (or plant parts) in resource-rich habitats. Evidence for and against both is available. Thus, gall-forming phylloxera (Box 11.2) prefers fast-growing meristematic tissue found in rapidly extending shoots of its healthy native vine host. In apparent contrast, the larva of Dioryctria albovitella (the pinyon pine cone and shoot boring moth; Pyralidae) attacks the growing shoots of nutrient-deprived and/or water-stressed pinyon pine (Pinus edulis) in preference to adjacent, less-stressed trees. Experimental alleviation of water stress has been shown to reduce rates of infestation, and enhance pine growth. Examination of a wide range of resource studies leads to the following partial explanation: boring and sucking insects seem to perform better on stressed plants, whereas gall inducers and chewing insects are adversely affected by plant stress. Additionally, performance of chewers may be reduced more on stressed, slow-growing plants than on stressed, fast growers.
The presence in Australia of a huge radiation of oecophorid moths whose larvae specialize in feeding on fallen eucalypt leaves suggests that even well-defended food resources can become available to the specialist herbivore. Evidently, no single hypothesis (model) of herbivory is consistent with all observed patterns of temporal and spatial variation within plant individuals, populations, and communities. However, all models of current herbivory theory make two assumptions, both of which are difficult to substantiate. These are:
- damage by herbivores is a dominant selective force on plant evolution;
- food quality has a dominant influence on the abundance of insects and the damage they cause.
Even the substantial evidence that hybrid plants may incur much greater damage from herbivores than either adjacent parental population is not unequivocal evidence of either assumption. Selection against hybrids clearly could affect plant evolution; but any such herbivore preference for hybrids would be expected to constrain rather than promote plant genetic diversification. The food quality of hybrids arguably is higher than that of the parental plants, as a result of less efficient chemical defenses and/or higher nutritive value of the genetically “impure” hybrids. It remains unclear whether the overall population abundance of herbivores is altered by the presence of hybrids (or by food quality per se) or merely is redistributed among the plants available. Furthermore, the role of natural enemies in regulating herbivore populations often is overlooked in studies of insect—plant interactions.
Many studies have demonstrated that phytophagous insects can impair plant growth, both in the short term and the long term. These observations have led to the suggestion that host-specific herbivores may affect the relative abundances of plant species by reducing the competitive abilities of host plants. The occurrence of induced defenses (Box 11.1) supports the idea that it is advantageous for plants to deter herbivores. In contrast with this view is the controversial hypothesis that “normal” levels of herbivory may be advantageous or selectively neutral to plants. Some degree of pruning, pollarding, or mowing may increase (or at least not reduce) overall plant reproductive success by altering growth form or longevity and thus lifetime seed set. The important evolutionary factor is lifetime reproductive success, although most assessments of herbivore effects on plants involve only measurements of plant production (biomass, leaf number, etc.).
A major problem with all herbivory theories is that they have been founded largely on studies of leaf- chewing insects, as the damage caused by these insects is easier to measure and factors involved in defoliation are more amenable to experimentation than for other types of herbivory. The effects of sap-sucking, leaf- mining, and gall-inducing insects may be as important although, except for some agricultural and horticultural pests such as aphids, they are generally poorly understood.
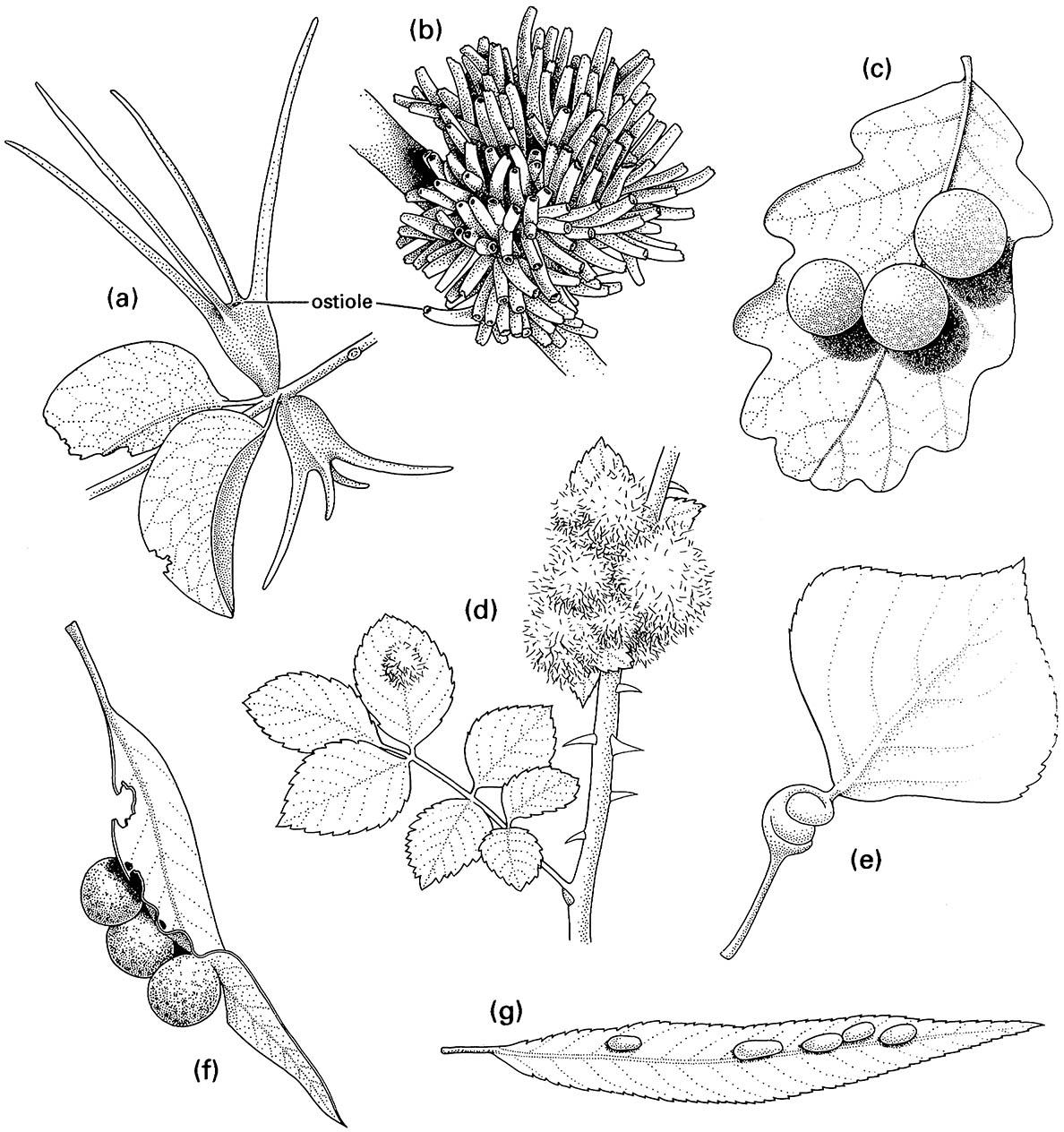
(a) two coccoid galls, each formed by a female of Apiomorpha munita (Hemiptera: Eriococcidae) on the stem of Eucalyptus melliodora; (b) a cluster of galls each containing a male of A. munita on E. melliodora; (c) three oak cynipid galls formed by Cynips quercusfolii (Hymenoptera: Cynipidae) on a leaf of Quercus sp.; (d) rose bedeguar galls formed by Diplolepis rosae (Hymenoptera: Cynipidae) on Rosa sp.; (e) a leaf petiole of lombardy poplar, Populus nigra, galled by the aphid Pemphigus spirothecae (Hemiptera: Aphididae); (f) three psyllid galls, each formed by a nymph of Glycaspis sp. (Hemiptera: Psyllidae) on a eucalypt leaf; (g) willow bean galls of the sawfly Pontania proxima (Hymenoptera: Tenthredinidae) on a leaf of Salix sp. ((d-g ) After Darlington 1975)

