11.4.1. Ant — plant interactions involving domatia
Domatia (little houses) may be hollow stems, tubers, swollen petioles, or thorns, which are used by ants either for feeding or as nest sites, or both. True domatia are cavities that form independently of ants, such as in plants grown in glasshouses from which ants are excluded. It may be difficult to recognize true domatia in the field because ants often take advantage of natural hollows and crevices such as tunnels bored by beetle or moth larvae. Plants with true domatia, called ant plants or myrmecophytes, often are trees, shrubs, or vines of the secondary regrowth or understorey of tropical lowland rainforest.
Ants benefit from association with myrmecophytes through provision of shelter for their nests and readily available food resources. Food comes either directly from the plant through food bodies or extrafloral nectaries (Fig. 11.10a), or indirectly via honeydew-excreting hemipterans living within the domatia (Fig. 11.10b). Food bodies are small nutritive nodules on the foliage or stems of ant plants. Extrafloral nectaries (EFNs) are glands that produce sugary secretions (possibly also containing amino acids) attractive to ants and other insects. Plants with EFNs often occur in temperate areas and lack domatia, for example many Australian Acacia species, whereas plants with food bodies nearly always have domatia, and some plants have both EFNs and food bodies. Many myrmecophytes, however, lack both of the latter structures and instead the ants “farm” soft scales or mealybugs (Coccoidea: Coccidae or Pseudococcidae) for their honeydew (sugary excreta derived from phloem on which they feed) and possibly cull them to obtain protein. Like EFNs and food bodies, coccoids can draw the ants into a closer relationship with the plant by providing a resource on that plant.
Obviously, myrmecophytes receive some benefits from ant occupancy of their domatia. The ants may provide protection from herbivores and plant competitors or supply nutrients to their host plant. Some ants aggressively defend their plant against grazing mammals, remove herbivorous insects, and prune or detach other plants, such as epiphytes and vines that grow on their host. This extremely aggressive tending is demonstrated by ants of Pseudomyrmex that protect Acacia in tropical America. Rather than protection, some myrmecophytes derive mineral nutrients and nitrogen from ant-colony waste via absorption through the inner surfaces of the domatia. Such plant “feeding” by ants, called myrmecotrophy, can be documented by following the fate of a radioactive label placed in ant prey. Prey is taken into the domatia, eaten, and the remains are discarded in refuse tunnels; the label ends up in the leaves of the plant. Myrmecotrophy occurs in the epiphytic Myrmecodia (Rubiaceae) (Fig. 11.11), species of which occur in the Malaysian and Australian regions, especially in New Guinea.
The majority of ant—plant associations may be opportunistic and unspecialized, although some tropical and subtropical ants (e.g. some Pseudomyrmex and Azteca species) are totally dependent on their particular host plants (e.g. Acacia or Triplaris and Cecropia species, respectively) for food and shelter. Likewise, if deprived of their attendant ants, these myrmecophytes decline. These relationships clearly are obligatorily mutualistic and, no doubt, others remain to be documented.
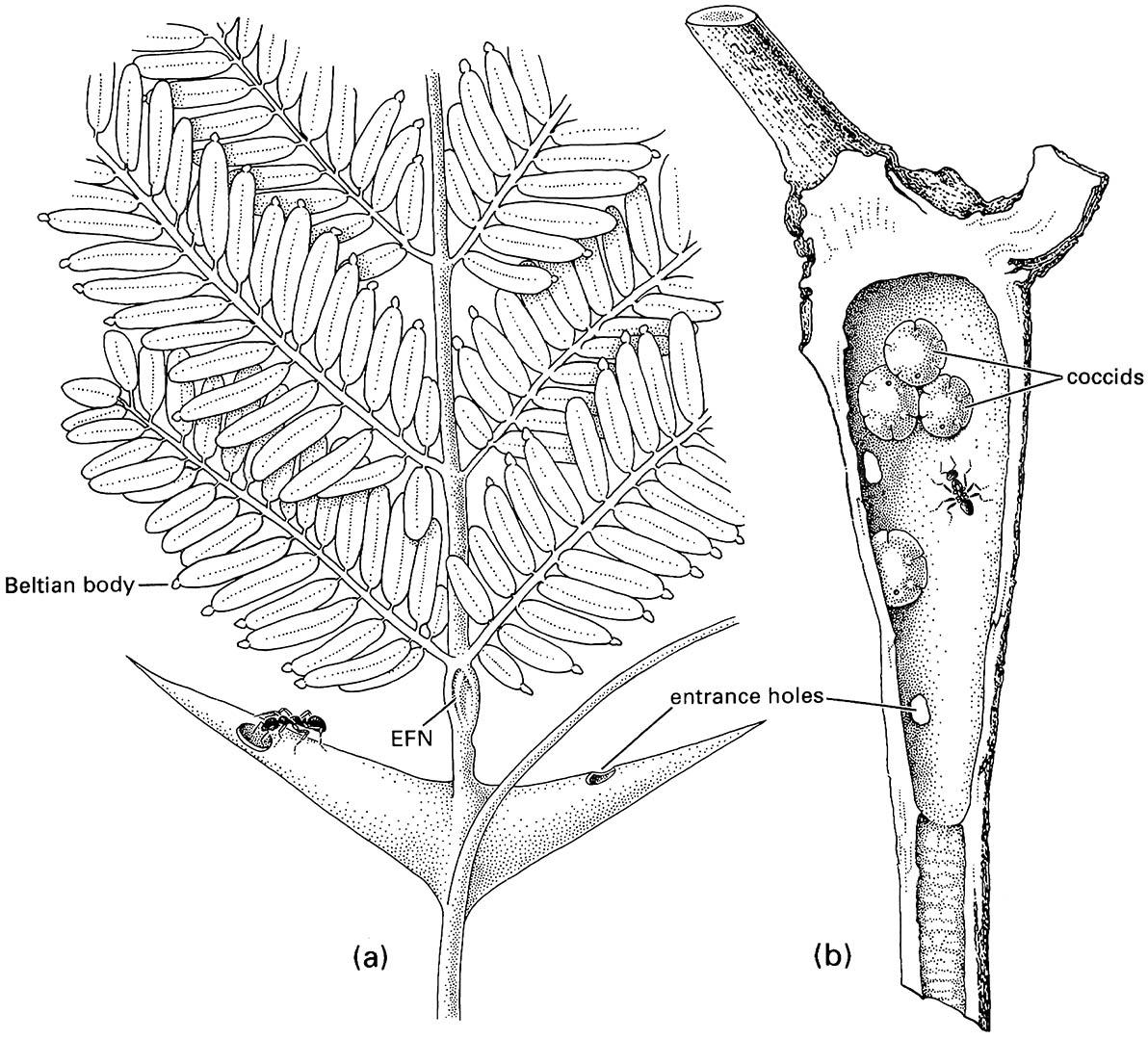
(a) a neotropical bull’s-horn acacia, Acacia sphaerocephala (Fabaceae), with hollow thorns, food bodies, and extrafloral nectaries (EFNs) that are used by the resident Pseudomyrmex ants; (b) a hollow swollen internode of Kibara (Monimiaceae) with scale insects of Myzolecanium kibarae (Hemiptera: Coccidae) that excrete honeydew that is eaten by the resident ants of Anonychomyrma scrutator. ((a) After Wheeler 1910; (b) after Beccari 1877)
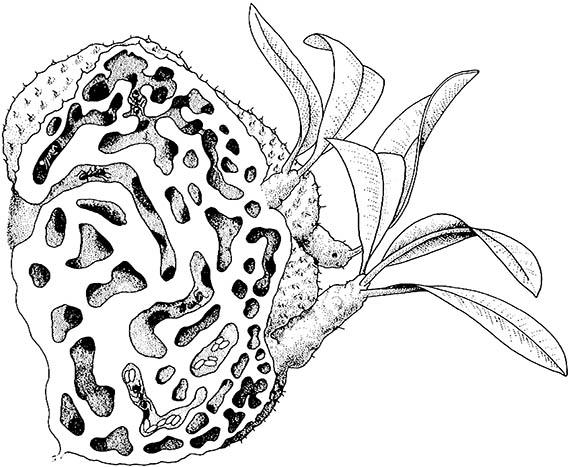
Ants live in smooth-walled chambers and deposit their refuse in warted tunnels, from which nutrients are absorbed by the plant. (After Monteith 1990)

