Эндокринная система
Эндокринная система насекомых образована разнородными нейросекреторными клетками, ретроцеребральным комплексом, проторакальной личиночной железой и нейрогемальными органами. Принимая участие в управлении метаболическими процессами, координируя и интегрируя деятельность разных систем организма, она подчиняется нервной системе и опосредует её функции на уровне гуморальных влияний. Вместе с тем эндокринная система управляет ходом развития и размножения насекомых.
Нейросекреторные клетки головного мозга
Отличаясь от нейронов способностью к синтезу гормонов, они рассредоточены по всем ганглиям нервной цепочки, образуя в головном мозге несколько обособленных групп. Различаясь по сродству к определённым красителям, эти клетки распределяются на четыре типа, обозначаемые А (А’, А’’) , В, С, D. Впрочем, далеко не все клетки одного типа выделяют секреты со сходными физиологическими свойствами. Клетки медиальной группы головного мозга, расположенные близ оцеллярных нервов, имеют аксоны, направленные к противоположным сторонам протоцеребрума (рис. 129), где проникают в первую пару кардиакальных нервов.
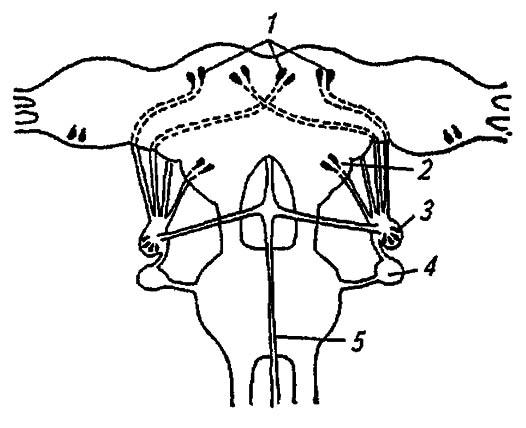
Рисунок 129. Нейросекреторная система головного мозга насекомых (по Тыщенко, 1977):
1 — медиальные и латеральные нейросекреторные клетки; 2 — тритоцеребрум; 3 — кардиальное тело; 4 — прилежащее тело; 5 — возвратный нерв
Выходя за пределы мозга, эти нервы направляются к кардиальным телам (corpora cardiaca), а иногда и к прилежащим телам (corpora allatum) ретроцеребрального комплекса. Группы латеральных нейросекреторных клеток формируют вторую пару кардиакальных нервов, а их третья и четвёртая пары образуются за счёт соответствующих клеток дейтоцеребрума и тритоцеребрума. Кроме того, нейросекреторные элементы обнаружены в оптических долях у некоторых насекомых. А- и В-клетки медиальных групп протоцеребрума саранчи Locusta migratoria L. синтезируют вещества, тормозящие деятельность прилежащих тел и влияющие на белковый обмен в гемолимфе, С-клетки — вещества, стимулирующие прилежащие тела и синтез меланинов, a D-клетки образуют диуретический гормон. Медиальные А’ - клетки тутового шелкопряда Bombux mori L. продуцируют активационный гормон, а В-клетки подглоточного ганглия — вещества, индуцирующие диапаузу.
Ретроцеребральный комплекс
Кардиальные тела помимо четырёх пар нервов, отходящих от головного мозга, имеют связи со стоматогастрической нервной системой и рассматриваются как её ганглии. Облечённые соединительнотканной оболочкой, они имеют собственные нейросекреторные элементы и нейроны, а также нейроглиальные клетки. Нередко кардиальные тела срастаются в непарный ганглий или (например, у мух) формируют вместе с прилежащими телами и проторакальной железой кольцевую железу. У таракана Periplaneta americana L. они выделяют активационный гормон (АГ), гипергликемический гормон, нейрогормон D, стимулирующий биение сердца, и вещество, влияющее на проведение нервного возбуждения, у бабочек-павлиноглазок — вещества, связанные с синтезом половых аттрактантов или стимулирующие откладку яиц.
Далеко не все эти гормоны и физиологически активные вещества синтезируются кардиальными телами, которые наряду с секреторными функциями исполняют роль нейрогемальных органов, то есть органов, запасающих нейросекреты головного мозга.
В частности, активационный гормон лишь накапливается в них, так же как и гормон, индуцирующий вылупление бабочек из куколки Кроме кардиальных тел к нейрогемальным органам относятся: аорта (у клопов), депонирующая нейросекреты в своих стенках; своеобразные перисимпатические образования, топографически связанные с системой непарного нерва; прилежащие тела. Расположенные позади кардиальных тел прилежащие тела накапливают поступающий из них нейросекрет, синтезируют и выделяют в гемолимфу ювенильный гормон (ЮГ), регулирующий ход развития и размножения, процессы метаболизма, изменения окраски покровов, интенсивность питания, а также индуцирующий диапаузу и влияющий на поведение. Прилежащие тела, связанные двумя парами аллатальных нервов с кардиальными телами, вместе с тем соединены нервами с подглоточным ганглием.
Парные проторакальные железы, расположенные обычно в грудном отделе и входящие у двукрылых в состав кольцевидной железы, имеются только у личинок. Дегенерируя при переходе организма во взрослое состояние, они сохраняются в течение всей жизни лишь у некоторых первичнобескрылых насекомых, подёнок, одиночных фаз саранчи и у рабочих особей термитов. Не связанные столь тесно, как компоненты ретроцеребрального комплекса, с нервной системой, проторакальные железы часто вообще не иннервируются. Однако посредством гуморальных влияний, распространяющихся по гемолимфе, они явно зависят от нейросекрета головного мозга, реагируя выделением линочного гормона — экдизона. По существу, проторакальные железы — компонент единой эндокринной системы насекомых, во многих отношениях напоминающей эндокринную систему позвоночных животных. При этом в отличие от позвоночных насекомые, по-видимому, лишены половых гормонов, определяющих развитие вторичных половых признаков, и лишь у жуков-светляков Lamipyris noctiluca L. при пересадке семенников в тело развивающейся личинки самки происходит реверсия пола и появляются признаки, свойственные самцу. Предположительно действующий в данном случае андрогенный гормон пока не выделен.
Имеются также данные о наличии в зрелых яичниках мухи Musca domestica L. оостатического гормона, который предотвращает развитие очередной партии яиц до тех пор, пока не будут отложены уже сформированные яйца. Этот гормон, синтезируемый в яичниках, тормозит выделение нейросекрета медиальными клетками головного мозга. В свою очередь, от этого нейросекрета зависит продукция ювенильного гормона, необходимого для стимуляции деятельности яичников. Близкий по характеру действия антигонадотропин клопа Rhodniusprolixus St., не влияя на другие эндокринные органы, блокирует активность самого ювенильного гормона непосредственно в яичниках.
Гормоны
У насекомых открыт целый- спектр гормонов разного действия, но далеко не все они идентифицированы. Активационный гормон (АГ), проявления которого были отмечены раньше других (в 1922 году, видимо, образован двумя компонентами: один из них стимулирует синтез РНК, другой влияет на проницаемость мембран и мембранный потенциал клеток проторакальной железы. Предположение об идентичности АГ холестерину не подтвердилось, но при выделении фракций нейросекрета медиальных клеток протоцеребрума удалось повысить их активность в 8 000 раз. При этом были выделены три основные фракции с разной молекулярной массой.
Воздействуя на проторакальные железы, АГ побуждает их к синтезу экдизона — гормона, обусловливающего наступление и прохождение линьки, а также склеротизацию покровов при совместном действии с бурсиконом. Имеются данные о возможности продукции экдизона гемоцитами, клетками жирового тела и эноцитами.
После того как Бутенандт и Карлсон, переработав в 1954 году 500 кг куколок тутового шелкопряда, получили 25 мг чистого кристаллического продукта, было показано, что он образован двумя стероидами: ?- и ?-экдизонами. Вопреки точке зрения на (3-экдизон как побочный продукт синтеза ?-экдизона имеются свидетельства о том, что ?-экдизон, синтезированный в проторакальных железах, — предшественник основного гормона (3-экдизона, синтез которого происходит в эноцитах и, возможно, в клетках жирового тела.
В настоящее время признан действительным следующий метаболический ряд: холестерин — 22, 25-деоксиэкдизон — 22-деоксиэкди-зон — ?-экдизон — (3-экдизон. Существенно, что оба экдизона и более 20 стероидов сходного физиологического действия выделены из растений, где их содержание достигает нередко очень высоких значений, например до 0,8 мкг на 1 г сухой массы в тканях тисса и ногоплодника. Однако, заимствуя стероиды из тканей кормовых растений, насекомые самостоятельно синтезируют холестерин и вещества всех последующих этапов метаболического пути экдизона.
Основными мишенями для экдизона служат клетки гиподермы и имагинальных дисков, точнее, хромосомы этих клеток. С позиций мембранной гипотезы экдизон, вызывая изменения проницаемости внутриклеточных мембран для ионов, стимулирует пуффинг — образование на хромосомах местных вздутий, соответствующих очагам интенсивного синтеза иРНК и считывания генетической информации. В политенных (многонитчатых) хромосомах слюнных желез двукрылых эти вздутия (пуфы) появляются в последовательности, соответствующей ходу линочного процесса. Введение экдизона провоцирует реализацию этой последовательности пуффинга на хромосомах даже взрослого насекомого.
В качестве альтернативы была выдвинута депрессорная гипотеза действия экдизона на хромосомы. Пробуждая опероны, ответственные за синтез специфических ферментов, экдизон в соответствии с указанной гипотезой непосредственно определяет последовательность считывания генетической информации.
Таким образом, экдизон обусловливает наступление и прохождение линочного процесса, однако его характер и результат зависят от ювенильного гормона, продуцируемого прилежащими телами. Химический анализ и синтез соединений, обладающих действием ЮГ, позволили выявить три близких терпеноида, условно названных ЮГ-I, ЮГ-II, ЮГ-III. Открыты и синтезированы также многие другие соединения, воспроизводящие эффекты этих терпеноидов. Названные аналогами ювенильного гормона, или ювеноидами, они были выделены из растений и животных или синтезированы искусственно.
Примечательно, что стимулом к поискам аналогов ЮГ послужили изготовленные из древесины пихты бальзамической картонные садки, в которых нимфы клопов-солдатиков (Pyrrhocoris apterus L.) не становились взрослыми, половозрелыми особями.
Среди ювеноидов особую известность приобрели близкие к гормонам насекомых синтетические нециклические сесквитерпены — фарнезол и его производные, а также моноциклические сесквитерпены типа ювабиона. В связи с выраженной специфичностью действия природные и синтетические ювеноиды используют для борьбы с отдельными видами вредителей сельскохозяйственных культур.
При наличии в гемолимфе ювенильного гормона линяющие насекомые испытывают лишь ограниченные преобразования, характерные, например, для перехода из одной преимагинальной стадии в другую.
Преобразования в имаго, особенно резкие при полном превращении, в присутствии ювенильного гормона становятся невозможными. Именно поэтому его иногда называют неотенином (гормоном молодых), или гормоном статус-кво.
В ходе развития к основным мишеням гормона относятся клетки гиподермы, а у вполне развитых самок — яичники. В соответствии с реверсивной гипотезой в клетках гиподермы насекомых присутствуют два или три типа генов (личиночные, куколочные и имагинальные), которые активируются при разных концентрациях гормона. С позиции репрессорной гипотезы действие ЮГ проявляется через подавление оперонов, ответственных за формирование имагинальных органов и структур. Однако возможность появления личиночных признаков у куколок большого мучного хрущака (реверсия) противоречит этому допущению. Вместе с тем ЮГ, по-видимому, способствует синтезу тРНК, которая устанавливает порядок в расположении аминокислот белков, синтезируемых на рибосомах.
Следовательно, гормоны насекомых выполняют многообразные функции — от регуляции активности генов до индукции диапаузы. Однако в первую очередь они ответственны за размножение и развитие, распространяя своё влияние вплоть до отдельных актов поведения, связанных с этими процессами. Например, ЮГ способен повышать двигательную активность самцов саранчовых или готовность к спариванию самок дрозофилы Drosophila melanogaster Mg. Экдизон, синтезируемый под воздействием созревающих в яичнике яиц, может блокировать стремление самок комара Anopheles freebomi Zt. к кровососанию, необходимому для формирования очередной партии яиц. Экстракты кардинальных тел, стимулируя фаллический нерв таракана Periplaneta americana L., обеспечивают реализацию стереотипа сексуального поведения.
В заключение отметим, что эндокринная система насекомых, координируя процессы метаболизма и ход развития, является системой долговременных процессов и реакций. В этом отношении она противопоставляется нервной системе, формирующей быстрые реакции на скоротечные стимулы. Кроме того, не имея собственных средств рецепции, эндокринная система теснейшим образом зависит от стимулов, опосредованных нервным возбуждением, и вместе с нервной системой обеспечивает интеграцию всех жизненно важных функций, направляя их на путь продолжения рода.

