Дыхание и газообмен
Снабжение органов и тканей кислородом достигается путём его диффузии через покровы тела и стенки трахей. Сходным образом из организма выводится углекислота и испаряется влага. Следовательно, при сравнительно высоком отношении площади поверхности тела к его объёму обеспечение интенсивного газообмена чревато постоянной угрозой иссушения. Чем меньше организм, тем это отношение больше, и поэтому только самые мелкие насекомые, обитающие во влажных средах или в воде, довольствуются «кожным дыханием».
Более крупные насекомые для увеличения поверхности диффузии газов и сокращения потерь воды развивают сложную систему внутренних тонкостенных трубочек — трахей, пронизывающих все тело и разветвляющихся в его полости.
Диффузия кислорода через воздух осуществляется в 3?105 быстрее, чем через воду, и в 106 раз быстрее, чем через ткани. Однако углекислый газ диффундирует через воду и ткани тела с гораздо большей лёгкостью, и уж если организм удовлетворяет свои потребности в кислороде, то он, несомненно, избавляется от углекислоты. Расчёты свидетельствуют о том, что трахейная система насекомых служит оптимальным средством решения проблем газообмена и предотвращения потерь воды.
Трахейная система.
Возникая в виде сегментарных впячиваний эктодермы, трахеи сформированных насекомых начинаются серией дыхальцевых отверстий, или стигм, открывающихся на плейритах средне- и заднегруди и восьми первых сегментах брюшка. Такая картина наблюдается далеко не всегда; чаще бывает сокращённый набор стигм, сохраняющихся, например, у личинок двукрылых лишь на среднегруди и на восьмом сегменте брюшка. Часто они совсем редуцируются, и трахейная система становится замкнутой.
Дыхальца бывают простыми или сложными. Простые дыхальца представляют собой отверстия, иногда снабжённые специальными замыкательными аппаратами в виде двух створок, как устьица на листьях растений. У большинства насекомых эти исходные отверстия погружаются под покровы, на которых развиваются новые, более сложные запирательные клапаны, ограничивающие доступ воздуха в расположенную под ними полость атриума (рис. 90). Последняя нередко заполнена густыми фильтрующими волосками или снабжена пористой пластинкой, предназначенной не столько для очистки воздуха от пылевидных частиц, сколько для предохранения трахей от заливания водой, проникновения в них паразитов, а также для сокращения потерь влаги, конденсирующейся на внутренней поверхности пластинки.

Рисунок 90. Строение стигмы трахейной системы (по Шванвичу, 1949):
1 — запирательный клапан; 2 — атриум
У примитивных первичнобескрылых насекомых от отверстий стигм отходят отдельные пучки древовидно-ветвящихся трахей, однако для насекомых более характерно объединение их в общие стволы.

Рисунок 91. Строение трахейной системы насекомых (по Romoser, 1981)
Весьма обычны идущие под покровами продольные боковые стволы, в которые открываются стигмы (рис. 91). Отходящие от них трахеи разделяются на все более тонкие веточки и постепенно переходят в трахеолы. Они проникают между отдельными органами и дольками жирового тела и укреплены спиральными утолщениями (тенидиями) общей кутикулярной выстилки — интимы (рис. 92). Благодаря этому они не спадают и сохраняют просвет при давлении и изгибаниях, как трубки противогаза.

Рисунок 92. Строение трахеи насекомых (по Romoser, 1981)
И тенидии, и ингима, секретируемые эпителием трахей, обычно не содержат хитина и воскового слоя эпикутикулы, но богаты эластичным резилином, не препятствующим газообмену. Как производные гиподермы они сбрасываются при линьках. Вытягиваемые тонкими трубочками через отверстия стигм, они замещаются вновь секретируемыми образованиями. Тенидии, отмеченные не только в трахеях, но и в трахеолах, не препятствуют их растяжению в длину, что имеет значение при постоянных движениях внутренних органов. При этом испытывающие кислородное голодание (гипоксию) клетки отсылают длинные псевдоподии к близлежащим трахеолам и подтягивают их к себе. Постоянный дефицит кислорода индуцирует развитие новых трахей и трахеол сразу же после линек.
Трахеолы, диаметр которых не превышает 1 мкм, не сбрасывают интимы в ходе линек. Они обычно заканчиваются в цитоплазме звездчатых клеток — трахеобластов (рис. 93), проникающих своими отростками вплоть до митохондрий клеток крыловых мышц, или плотно прилегают к их оболочкам снаружи.

Рисунок 93. Ветвление трахеолы в трахеобласте (по Romoser, 1981):
1 — трахесшы; 2 — трахеобласт
Общее число трахеол огромно и у крупных активных насекомых достигает (1—1,5)х106. Некоторые крупные стволы теряют тенидии и преобразуются в объемистые воздушные мешки, способные сплющиваться и раздуваться. Запасая воздух, содействуя циркуляции гемолимфы и сохранению тепла, они амортизируют резкие биения крыловых мышц и сокращают удельный вес тела при полете. У некоторых насекомых такие мешки выполняют роль резонаторов в звуковых органах.
Легко проникая сквозь тонкие стенки трахей и трахеол, кислород растворяется в полостной жидкости и диффундирует на расстояние до 1 мм. Однако для обеспечения работы крыловых мышц рассчитанные расстояния до трахеол должны быть сокращены в сотни раз (до 6–8 мкм). На самом деле они не превышают 2–3 мкм.
Если в гемолимфе имеется гемоглобин, то он связывается с кислородом в оксигемоглобин и транспортирует кислород по тканям, но только при его дефиците в окружающей среде. Это имеет определённое значение для обитающих в гниющем иле личинок комаров-звонцов, личинок оводов, паразитирующих в кишечнике млекопитающих, или для клопов-гладышей, надолго погружающихся под воду. В противном случае оксигемоглобин лишь резервирует запасы кислорода и не участвует в его транспортировке, а газообмен осуществляется обычными для насекомых способами. Что касается углекислоты, то, диффундируя по тканям в 35 раз быстрее кислорода, она легко выводится из организма через покровы и стенки трахей.
Расчёты размеров трахейной системы и потребностей насекомого в кислороде показали, что уже двухпроцентный перепад его концентраций в тканях тела и в атмосфере достаточен для диффузии. Таким образом, трахейная система насекомого с открытыми стигмами вмещает почти десятикратный избыток кислорода, но при полете его расходование возрастает в 50 раз и более. К тому же при открытых стигмах резко возрастают транспирация и утечка воды из организма. По-видимому, именно эти обстоятельства определили развитие специальных средств пассивной и активной вентиляции трахейной системы у крупных активных насекомых.
Пассивная вентиляция обеспечивается расходованием кислорода и растворением углекислоты в гемолимфе. Возникающий вследствие этого вакуум приводит к засасыванию воздуха через плотно сомкнутые створки стигм. Накопление углекислого газа требует его выведения, которое у некоторых насекомых приобретает характер периодически повторяющихся «залпов» (рис. 94). Например, у диапаузирующих куколок шелкопряда Hyalophora cecropia L. через каждые 7 часов створки дыхалец раскрываются и углекислота, легко диффундирующая из трахей и гемолимфы, выводится наружу за несколько минут. Между тем потребление кислорода происходит непрерывно, а смыкание дыхалец в интервалах между «залпами» сокращает транспирацию воды. Сходные явления отмечены у личинок комаров, периодически всплывающих на поверхность воды за очередной порцией воздуха.
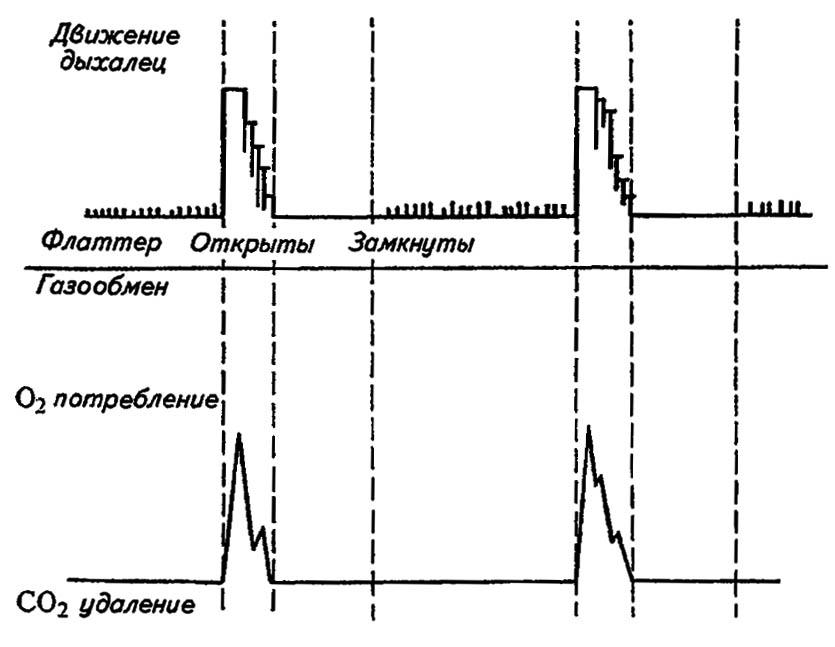
Рисунок 94. Диаграмма работы дыхалец, демонстрирующая «залпы» выведения углекислоты (по Gillot, 1980)
Весьма характерны средства пассивной вентиляции у обитателей водоёмов и у внутренних паразитов. Рассмотренное выше строение трахейных жабр личинок подёнок и стрекоз допускает диффузию растворенных в воде газов. Проникая в омывающую изнутри жабры гемолимфу, кислород переходит в трахеи и трахеолы. Кутикулярные (дыхальцевые) жабры (рис. 95), свойственные водным куколкам некоторых двукрылых, обеспечивают ещё более лёгкий газообмен.

Рисунок 95. Кутикулярные жабры личинки подёнки (по Шванвичу, 1949)
Связанные непосредственно со стигмами как выступающие из них тонкие кутикулярные пузыри, они не требуют предварительного растворения кислорода в гемолимфе и вместе с тем легко выводят углекислоту. Сходным образом функционируют пластроны — воздухоносные полости в гидрофобных сеточках и решётках на покровах (рис. 96). Являясь органами газообмена многих водных насекомых или их яиц, они способны обеспечивать и диффузию газов из атмосферы.

Рисунок 96. Пластрон яиц насекомых (по King, 1982):
а и б — разрезы на разных уровнях пластрона
Многие водные насекомые все же дышат кислородом атмосферы, поднимаясь время от времени на поверхность. Часто они уносят с собой в глубину пузырьки воздуха. За счёт диффузии газов в погруженный под воду пузырек воздуха обновляемые в нем запасы кислорода позволяют этим насекомым долгое время оставаться под водой. Другие насекомые используют кислород воздухоносных полостей подводных растений или развивают различные сифоны и дыхательные трубки. Приспосабливаются к дыханию кислородом атмосферы и некоторые эндопаразиты, либо выставляя наружу дыхательные рожки, либо прикрепляясь к воздушным мешкам и к стволам трахейной системы хозяина. Однако при этом они уже используют средства активной вентиляции трахей.
Активная вентиляция обеспечивается регулярными движениями тела, имитирующими «вдохи» и «выдохи», и согласованной работой дыхалец. Попеременно смыкаясь и раскрываясь, передние и задние дыхальца недолго остаются открытыми. Во время активного полёта грудные дыхальца вообще не закрываются, а за счёт сокращений мощных крыловых мышц создаётся автономная вентиляция грудного отдела.
Активная вентиляция обеспечивает «проветривание» только главных трахейных стволов и не способна усилить потребление кислорода клетками активно работающих органов. Для этих целей служит «трахеолярный контроль газообмена», суть которого в следующем. У насекомого, не занятого активной деятельностью, концы трахеол заполнены жидкостью и выключены из распределения кислорода или поддерживают весьма слабую диффузию газов. При активной работе накапливающиеся в гемолимфе метаболиты повышают осмотическое давление и жидкость, заполняющая окончания трахеол, легко проходит через их стенки. Сразу же заполняемые воздухом, они существенно увеличивают поверхность активного газообмена. Последующее окисление и выведение метаболитов содействуют падению осмотического давления гемолимфы, и её плазма устремляется в тонкие капилляры трахеол до тех пор, пока свойственные им капиллярные силы не уравновесят осмотические.
Недостаток кислорода или избыток углекислоты приводит к усилению дыхательных движений, изменению ритма работы стигм и включению некоторых дополнительных механизмов вентиляции (рис. 97). Координированное сокращение мышц тела при дыхательных движениях определяется импульсами, поступающими от мета-мерно расположенных первичных нервных центров дыхания. Вторичные центры, находящиеся в грудных и головных ганглиях, стимулируют или тормозят деятельность первичных центров. Регуляция работы дыхалец обеспечивается иннервацией мускулатуры створок и существенным образом зависит от влажности воздуха и концентрации ионов в гемолимфе, регистрируемой специальными центрами головного мозга.

Рисунок 97. Схема регуляции дыхания у насекомых (по Тыщенко, 1976)
Газообмен.
Потребление кислорода и выделение углекислоты — внешние проявления газообмена, суть которого состоит в окислении энергетических субстратов (образовавшихся при переваривании углеводов, жирных кислот и аминокислот). При окислении углеводов объем выделенной углекислоты равен объёму поглощённого кислорода и их отношение — дыхательный коэффициент (объем CO2 /объем O2) — равен 1. При окислении продуктов расщепления жиров и белков этот коэффициент оказывается несколько меньшим (для жиров 0,77–0,82, белков 0,7) и, очевидно, по его значению можно судить о характере используемой пищи. Если он особенно мал (менее 0,7), то некоторая часть жиров, по-видимому, расходуется на синтез углеводов, и, наоборот, при синтезе жиров из углеводов дыхательный коэффициент превышает 1.
С потребления кислорода начинаются реакции, в результате которых накапливается энергия в макроэргических связях АТФ (в меньшей степени молекул аргининфосфата и ацетилкофермента А) и выделяются конечные продукты обмена — CO2 и H2O.
Освобождение энергии обычно происходит двумя путями. При гликолизе — анаэробном распаде углеводов в цитоплазме — у позвоночных животных образуется молочная кислота, у насекомых наряду с ней — пировиноградная кислота и проникающий через мембраны митохондрий ?-глицерофосфат (рис. 98). При окислительном фосфорилировании — аэробном окислении жирных кислот, углеводов и аминокислот в митохондриях — процессы распада углеводов протекают либо в пентозном цикле, либо в цикле Кребса. В первом случае, испытав ряд преобразований, молекула глюкозы даёт начало молекуле триозы и трём молекулам углекислоты.

Рисунок 98. Пути гликолиза у насекомых (по Тыщенко, 1976)
Значение пентозного цикла не исчерпывается освобождением энергии. Например, у таракана Periplaneta americana L. при этом используется только 49 % глюкозы. Однако в пентозном цикле синтезируются пенгозы, входящие в состав нуклеотидов и нуклеиновых кислот, что определяет реальное значение пентозного цикла в метаболизме насекомых.
Таким образом, основная масса углеводов подвергается распаду в ходе гликолиза либо окисляется в цикле Кребса, который начинается с продуктов превращения пировиноградной кислоты (рис. 99). Эта кислота включается в цикл либо через образование щавелево-уксусной кислоты, что, однако, отмечено только у мухи цеце, либо через ацетилкофермент А. Далее она претерпевает ряд превращений, начиная с лимонной кислоты и заканчивая вновь щавелево-уксусной. Последняя связывается с ацетилкоферментом А и, давая начало лимонной кислоте, обеспечивает повторение цикла. В результате всех этих превращений образуются конечные продукты метаболизма: CO2 и H2O и макроэргические связи АТФ.
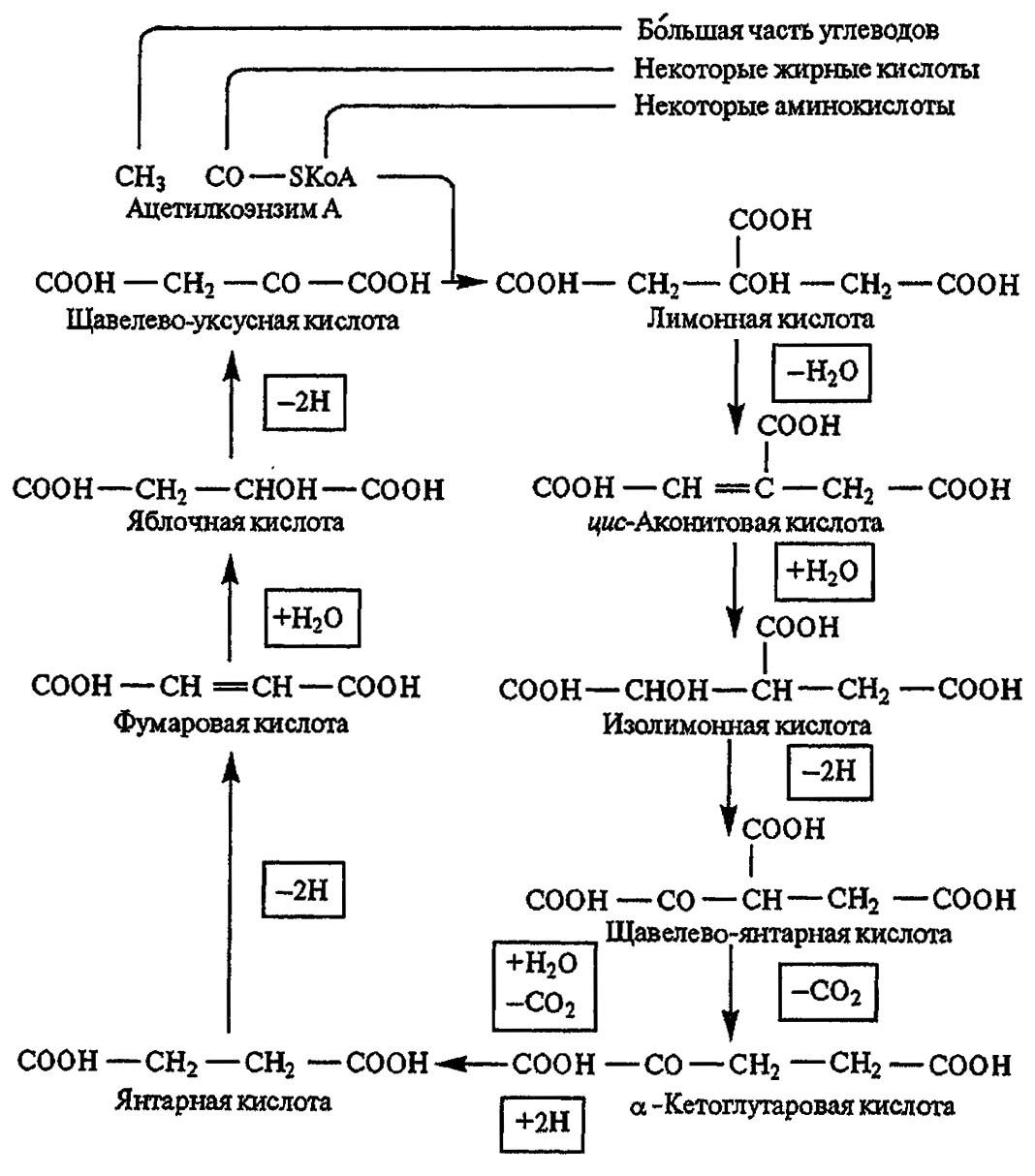
Рисунок 99. Основные звенья цикла Кребса (по Тыщенко, 1976)
Если в процессе гликолиза одна молекула глюкозы даёт начало двум макроэргическим связям, то в цикле Кребса формируется 38 связей. Сопоставление этих цифр не требует комментариев. Ферменты, катализирующие реакции цикла Кребса у насекомых, гораздо активнее, чем у позвоночных животных.
Наряду с углеводами окислительному фосфорилированию подвергаются жирные кислоты и аминокислоты, причём последние, вступая в цикл Кребса, испытывают трансаминирование. В результате формируются новые аминокислоты и аммиак, который связывается с образовавшейся при гликолизе пировиноградной кислотой.
Следовательно, в цикле Кребса скрещиваются важнейшие пути метаболизма белков, жиров и углеводов — всех основных компонентов перевариваемой в кишечнике пищи. В конечном счёте от них остаются вода, углекислый газ, подлежащие выведению экскременты, синтезированные новые органические соединения и запасы энергии.
Отметим, что некоторые дыхательные яды, например оксид углерода или производные синильной кислоты, блокируют дыхательные ферменты. Однако при пониженном газообмене зимующих (диапаузирующих) насекомых, который обеспечивается совершенно особыми, специфическими ферментами, эти яды не оказывают действия даже в концентрациях, смертельных при активной жизнедеятельности.

