Нервная система
Основной структурный и рабочий элемент нервной системы — нейрон. Это специализированная нервная клетка с собственной оболочкой, набором внутриклеточных органелл и нейрофибриллами (рис. 111). От её тела отходят длинный осевой отросток — аксон и короткие ветвящиеся дендриты. Получая нервные импульсы от других нейронов, дендриты переводят их на аксон, по которому возбуждение распространяется без затухания на другие нейроны или эффекторы — разного рода железы и мышцы.
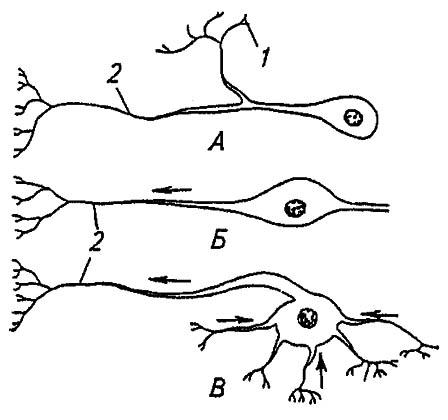
Рисунок 111. Типы нейронов насекомых (по Gillot, 1980):
А, Б, В — соответственно униполярный, биполярный, мультиполярный нейрон; 1 — дендриты; 2 — аксон
В зависимости от исполняемых функций нейроны подразделяются на сенсорные (чувствительные), проводящие возбуждение от рецепторов к нервным центрам; ассоциативные (вставочные, или интернейроны), перерабатывающие поступающую в центры информацию; моторные (двигательные) нейроны, которые доносят в центробежном направлении до эффекторов возбуждение, полученное от ассоциативных нейронов (рис. 112). Сформированная таким образом трёхнейронная рефлекторная дуга обеспечивает целесообразное реагирование на стимулы, тогда как образованная только сенсорным и моторным нейронами двухнейронная дуга обусловливает однозначный ответ на раздражение. Свойственная более примитивным организмам, она не характерна для насекомых, рефлекторные дуги которых нередко включают по нескольку ассоциативных нейронов, обеспечивая тем самым весьма сложные формы реагирования.
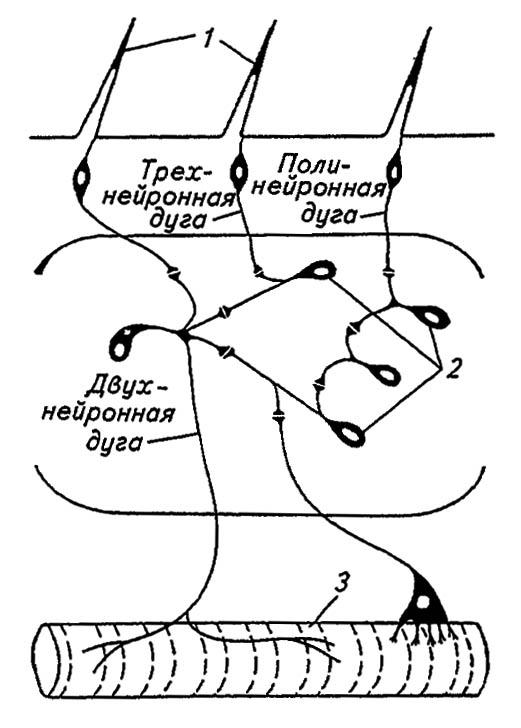
Рисунок 112. Рефлекторные дуги насекомых (по Тыщенко, 1977):
1 — рецепторы; 2 — ассоциативные нейроны; 3 — эффектор
В соответствии с предназначением сенсорные нейроны обычно являются биполярными, так как на полюсе, противоположном аксону, имеют единственный дендрит; реже они несут несколько дендритов и становятся мультиполярными (см. рис. 111). Тела этих нейронов расположены на периферии и входят в органы чувств,, тогда как тела моторных нейронов сосредоточены в ганглиях (нервных узлах) нервной системы, а отходящие от них аксоны выходят на периферию. Как правило, моторные нейроны униполярны: и дендрит, и аксон отходят от общего корня с одной стороны клетки. Распространяющиеся по ним импульсы нервного возбуждения достигают мышечных волокон или иных эффекторов. Ассоциативные нейроны биполярны, и их аксоны, изредка достигающие нескольких сантиметров в длину («гигантские аксоны»), проходят по ганглиям нервной системы.
Взаимоотношения разных нейронов в нервных центрах показаны на схеме рефлекторных дуг (см. рис. 112), а их взаимное положение — на поперечном срезе одного из ганглиев брюшной нервной цепочки (рис. 113). Облечённый соединительнотканной оболочкой (нейрилемма) и подстилающим слоем клеток (перинейрум), каждый ганглий прямокрылых насекомых имеет дополнительную защиту от контакта с гемолимфой в виде слоя клеток жирового тела и эпителия трахей. Клетки перинейрума регулируют транспорт ионов через нейрилемму и, поддерживая постоянство ионного состава, обеспечивают проведение нервного возбуждения без внешних помех. Эти клетки также переводят из гемолимфы необходимые для нейронов вещества, передавая их внутриганглионарным (нейроглиальным) клеткам. Последние через цитоплазматическне выросты питают нейроны или образуют вокруг их аксонов плотные спирально закрученные оболочки со свойствами изоляторов.
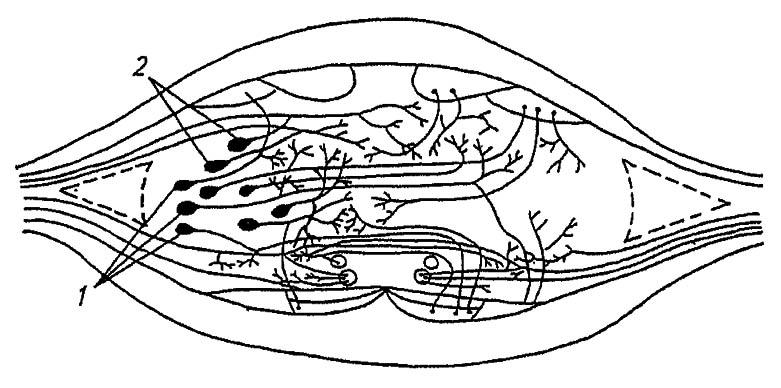
Рисунок 113. Поперечный срез через ганглий брюшной нервной цепочки. Тела ассоциативных (1) и моторных (2) нейронов (по Шванвичу, 1949)
Нейроны с обслуживающими их нейроглиальными клетками распределены во внешнем, корковом, слое ганглия, непосредственно под перинейрумом, тогда как центральная часть ганглия — нейропиль — занята сплетенными нервными волокнами. На периферии нейропиля с дорзальной стороны проходят аксоны моторных нейронов, формируя выходящие из ганглия дорзальные корешки боковых нервов. С вентральной стороны в нейропиле сосредоточены аксоны сенсорных нейронов, входящие в ганглий через вентральные корешки боковых нервов, а центр нейропиля заполнен разветвлениями ассоциативных нервных клеток.
Сквозь нейропиль проходят также пучки нервных волокон, связывающих нейроны обеих сторон одного ганглия и соседних ганглиев друг с другом. Общее число нейронов в пределах одного ганглия обычно не превышает 1 000, а в пределах всей нервной системы насекомого измеряется несколькими миллионами. Это весьма малые величины по сравнению с таковыми у млекопитающих, однако сложность реакций и форм поведения насекомых свидетельствует о совершенстве их нервной системы.
Центральная нервная система
Она образована двойной цепью ганглиев, связанных между собой продольными коннективами и поперечными комиссурами (рис. 114). Её передний отдел — надглоточный ганглий, по положению и функциям отвечающий головному мозгу позвоночных животных, происходит из слившихся между собой ганглиев по меньшей мере трёх сегментов. Вслед за ним расположен под-глоточный ганглий, связанный с головным мозгом окологлоточными коннективами. В его состав входят соединённые друг с другом ганглии трёх сегментов челюстей. Далее следуют три грудных ганглия, изредка образующие единую ганглиозную массу, и, наконец, ганглии брюшных сегментов. Закладывающиеся у эмбриона в виде сегментарных зачатков, они позднее смещаются вперёд и нередко объединяются. У имаго наиболее примитивных насекомых (Thysanura) сохраняется не более восьми парных ганглиев брюшка, причём последний, самый крупный из них, имеет следы объединения 2–3 ганглиев.
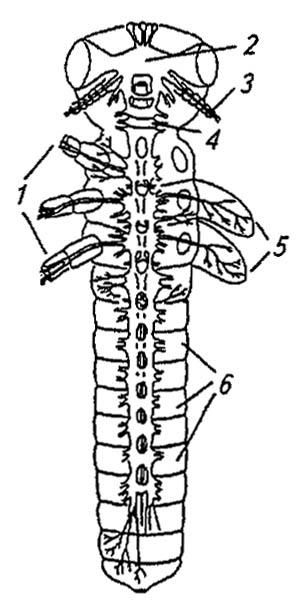
Рисунок 114. Центральная нервная система насекомых (по Шванвичу, 1949):
1 — ноги; 2 — надглоточный ганглий; 3 — антенна; 4 — подглоточный ганглий; 5 — крылья; 6 — ганглии брюшной нервной цепочки
У нимф подёнок и стрекоз в брюшке отмечено семь парных ганглиев, у прямокрылых — пять, другие насекомые имеют часто ещё большую концентрацию брюшной нервной цепочки. Например, центральная нервная система высших мух представлена лишь двумя ганглиозными массами (синганглиями): одна из них — головной мозг, другая — продукт объединения остальных ганглиев. Обычно личинки имеют менее концентрированную нервную систему, чем имаго, но у личинок мух единая ганглиозная масса вообще не разделена (рис. 115).
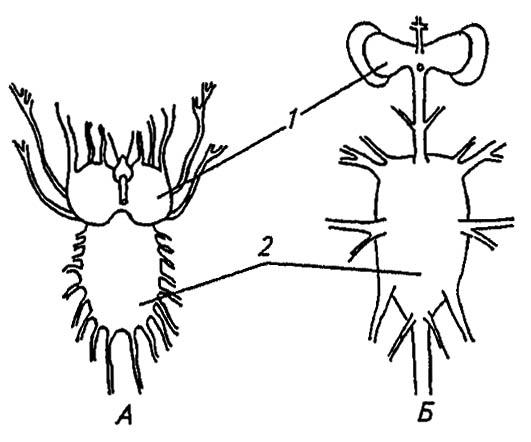
Рисунок 115. Центральная нервная система личинки и имаго мух:
А — личинка; Б — имаго; 1, 2 — соответственно головной и туловищный синганглий
Строение ганглиев брюшной нервной цепочки и ганглиев подглоточного нервного узла сходно; различия между ними определяются различиями иннервируемых органов. В частности, самый последний ганглий контролирует спаривание и откладку яиц и, как и впереди лежащие ганглии, имеет по две пары нервов, включающих сенсорные и моторные волокна. В ганглии крылоносных сегментов входят три пары нервов, обслуживающих ноги, крылья и туловищную мускулатуру, а в ганглий лишённой крыльев переднегруди и в ганглиозные массы подглоточного нервного узла — две пары нервов.
Головной мозг
Особой сложности в строении и во взаимодействии отдельных нервных центров достигает надглоточный ганглий (рис. 116). Образованный протоцеребрумом, дейтоцеребрумом и тритоцеребрумом, он включает ассоциативные центры и ганглиозные массы объединённых сегментов головы.
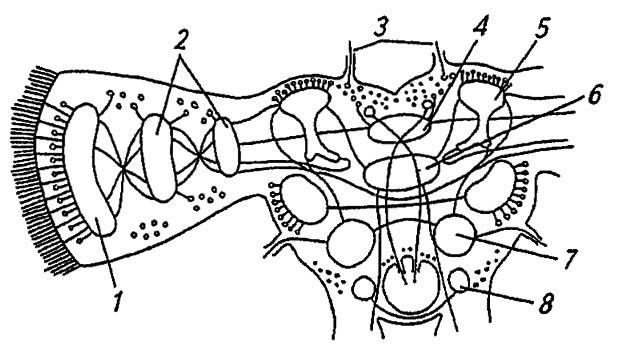
Рисунок 116. Головной мозг насекомых (по Romoser, 1981):
1 — оптическая пластинка; 2 — медуллярные пластинки; 3 — оцеллярные нервы; 4 — протоцеребральный мост; 5, 6 —соответственно грибовидное и центральное тело; 7 — дейтоцеребрум; 8 — тритоцеребрум
Более крупный протоцеребрум обладает явственными оптическими долями (с тремя парами нейропилярных масс и оцеллярным центром), воспринимающими и перерабатывающими сенсорную информацию от сложных глаз и дорзальных глазков насекомого. При этом тела сенсорных нейронов глазков связываются с оцеллярными ганглиями, от которых отходят оцеллярные нервы к межцеребральной части головного мозга. Нейропилярные, то есть образованные отростками нервных клеток, массы протоцеребрального моста и центрального тела соединяют симметричные доли протоцеребрума и, воспринимая аксоны от других центров, координируют моторную активность сегментов тела, определяя, в частности, согласованность работы дыхалец. Примыкающие с обеих сторон к долям протоцеребрума парные стебельчатые (или грибовидные) тела являются высшим ассоциативным центром, где замыкаются условно-рефлекторные связи. Их разрушение ведёт к утрате приобретенных навыков, но безусловные рефлексы сохраняются.
В чашечках стебельчатых тел расположены многочисленные (например, у тараканов до 300 000–400 000) ассоциативные нейроны, аксоны которых проходят в два стебелька, где образуют контакты с аксонами других центров протоцеребрума. Один из этих стебельков (а-доля) связывается с сенсорными, второй (?-доля) — с моторными нейронами (рис. 117). Степень развития стебельчатых тел коррелирует со сложностью поведения насекомых. Например, у муравьёв эти тела занимают 1/5 объёма мозга, а у рабочих пчёл они развиты значительно сильнее, чем у маток или трутней.
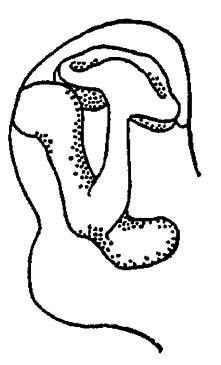
Рисунок 117. Грибовидное (стебельчатое) тело головного мозга насекомых (по Тыщенко, 1977)
Следующий отдел головного мозга — дейтоцеребрум — образован парными антеннальными долями, связанными друг с другом и с сенсорными и моторными аксонами антенн. Отходящий от них ольфакторный тракт достигает чашечек стебельчатых тел, обеспечивая участие ольфакторных стимулов в общей координации нервной деятельности.
Наконец, самый задний отдел головного мозга — крошечный тритоцеребрум — иннервирует головную капсулу с верхней губой и соединяется со стоматогастрической нервной системой (см. ниже) посредством фронтального ганглия и с брюшной нервной цепочкой посредством окологлоточного нервного кольца. Связывающие волокна (поперечная комиссура) между правой и левой долями тритоцеребрума проходят позади рта. Это свидетельствует об исходной принадлежности тритоцеребрума к туловищным сегментам насекомых, лишь вторично вошедшим в состав головы.
Завершая обсуждение строения центральной нервной системы, отметим, что подглоточный ганглий, обслуживающий рецепторы и мышцы челюстей и шейной области, содействует поддержанию двигательной активности насекомого. В этом смысле он служит антагонистом тормозящих влияний головного мозга.
Симпатическая нервная система
Именуемая также висцеральной или вегетативной, она слагается из стоматогастрической (ротожелудочной) системы, системы непарного нерва и каудальной системы (рис. 118). Её высший нервный центр — тритоцеребрум, от которого отходят два фронтальных коннектива к непарному фронтальному ганглию. Образуемый из стенки передней кишки эмбриона, этот ганглий вскоре теряет с ней связь и, соединяясь с головным мозгом, отсылает пару нервов к верхней губе и непарный возвратный нерв к гипоцеребральному ганглию. Последний связывается с нейросекреторными органами ретроцеребрального комплекса и с двумя вентрикулярными ганглиями. Иннервируя передние отделы кишечника и аорту, ганглии стоматогастрической системы включают в себя не только моторные и ассоциативные нейроны, но и сенсорные. В этом отношении они противопоставляются ганглиям брюшной нервной цепочки, с которыми связана система непарного нерва.
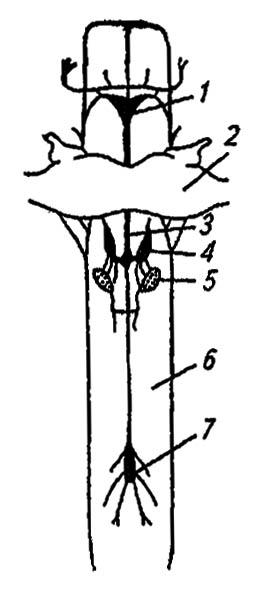
Рисунок 118. Симпатическая нервная система насекомых (по Шванвичу, 1949):
1 — фронтальный ганглий; 2 — мозг; 3 — возвратный нерв; 4 — затылочный ганглий; 5 — прилежащие тела; 6 — пищевод; 7 — вентрикулярный ганглий
Обслуживая дыхальца, эта система связана с ними боковыми ветвями, но образующие их нейроны находятся в грудных и брюшных ганглиях центральной нервной системы. Непарный нерв, по-видимому, участвует в работе крыловых мышц, так как при его разрушении резко возрастает их утомляемость.
Каудальная система, образованная двумя нервами, отходящими от последнего ганглия брюшка, иннервирует заднюю кишку и органы размножения насекомых.
Периферическая нервная система
Все нервы, выходящие из ганглиев центральной и симпатической нервной системы, наряду с сенсорными нейронами органов чувств (рецепторов) образуют периферическую нервную систему насекомых. Некоторые её компоненты будут рассмотрены позднее в связи с сенсорными системами и рецепцией.
Проведение нервного возбуждения в ганглиях
При прохождении возбуждения через ганглий возникают дополнительные ограничения, связанные с необходимостью преодоления по крайней мере нескольких синапсов рефлекторных дуг.
Отличаясь односторонней проницаемостью, синапсы допускают распространение импульсов только в одном направлении — от сенсорных нейронов к моторным, а необходимость выделения медиаторов и их диффузии по синаптической щели всякий раз задерживает импульс на 1–4 мс. Эта задержка многократно возрастает в полинейронных рефлекторных дугах со многими нейронами и синапсами. Вместе с тем нередкое для насекомых объединение множества пресинаптических волокон в одно постсинаптическое проявляется в так называемой пространственной суммации возбуждения; лишь совместное одновременное раздражение нескольких из них генерирует потенциал действия (ПД) в постсинаптическом аксоне. Аналогичным образом слишком слабые, но многократные раздражения, чередуясь в определённом ритме, приводят к временной суммации раздражений и к преодолению синаптического барьера.
Отметим также трансформацию ритма возбуждения во многих синапсах ганглия, своеобразный эффект последействия, связанный с продолжающейся после выключения раздражителя генерацией импульсов, и способность к иррадиации возбуждения на другие рефлекторные дуги ганглия. Наряду с привыканием (то есть блокированием синапса в ответ на монотонные раздражения) и обучением (облегчением переноса возбуждения через синапс) эти свойства ганглиев и синапсов приводят к фильтрации и преобразованию информации, которая поступает в нервные центры от рецепторов. Все это определяет целесообразность реагирования и сложность форм поведения насекомых.
Медиаторы нервного возбуждения в нервно-мышечных контактах и в синапсах ганглиев различны. Есть веские основания считать, что здесь, как и у позвоночных животных, медиатором служит ацетилхолин, разрушаемый ацетилхолинэстеразой. В частности, среди веществ, блокирующих активность ацетилхолинэстеразы насекомых, основное значение имеют фосфорорганические инсектициды.
Не касаясь более сложных аспектов функционирования центральной нервной системы насекомых, укажем, что каждый ганглий брюшной нервной цепочки — первичный рефлекторный центр иннервируемого им сегмента. Однако роль вторичных рефлекторных центров, контролирующих частные функции отдельных ганглиев, принадлежит головному мозгу и подглоточному нервному узлу. Комбинирование сегментарных рефлексов в целостные акты поведения возможно благодаря неспецифическим системам торможения и возбуждения, сравнимым с соответствующими системами млекопитающих.
Первая из них связана с нейронами стебельчатых тел протоцереребрума, вторая — с его центральным телом и с подглоточным ганглием. Именно поэтому лишённое мозга насекомое проявляет расторможенную двигательную активность. Вместе с тем в некоторых нейронах протоцеребрума наблюдается регулярная изменчивость в спонтанной генерации импульсов возбуждения. Являясь своеобразными «датчиками времени» и определяя сроки торможения и возбуждения двигательных центров, они тем самым контролируют суточную ритмику активности насекомого, которая сохраняется при постоянной темноте или при постоянном освещении.
Головной мозг насекомых исполняет роль высшего анализатора. Принимая участие в анализе сигналов, поступающих от всех органов чувств, он синтезирует акты поведения целостного организма. Лишённые мозга насекомые ведут себя как автоматы с испорченной программой: спонтанные рефлексы теряют приспособительный смысл и противоречат друг другу.
Многие аспекты высшей нервной деятельности насекомых ещё не изучены, но не следует думать, что существование и поведение насекомых целиком подчинены безусловным рефлексам, предопределяющим стереотипные реакции на все стимулы окружающего мира. Некоторые виды (например, пчела) способны к обобщению зрительных образов, превосходя в этом отношении рыб и даже крыс.

