Питание и переваривание пищи
Пищевая специализация. Подавляющее большинство насекомых — фитофаги, то есть потребители органического вещества, синтезированного и накопленного растениями. Однако среди них нередко встречаются хищники, паразиты, а также сапрофаги, питающиеся гниющими органическими остатками, некрофаги (поедают трупы) и копрофаги (источник пищи — навоз).
Широта спектра избираемой пищи предполагает разнообразие способов её отыскания и регистрации, морфо-функциональное многообразие пищеварительных органов и хода метаболизма. Лишь немногие насекомые всеядны (пантофаги) и одинаково охотно используют в пищу и растения, и животных, и перегнивающие остатки. Более распространены многоядные формы (полифаги), избирающие для себя лишь растения или животных, однако в основной массе насекомые относятся к олигофагам, например питаются растениями одного или нескольких близких семейств, проявляя явную избирательность и пищевую специализацию.
Среди паразитов нередки олигофаги, но среди хищников все же преобладают полифаги. Предел специализации — монофагия, то есть способность питаться организмами одного или немногих близких видов. Среди фитофагов монофагами являются многие тли, червецы и щитовки, среди паразитов — некоторые наездники, вши, блохи, среди хищников — божья коровка Rodolia cardinalis Muls., питающаяся исключительно австралийским желобчатым червецом Icerya purchasi Mask.
Кроме функции снабжения организма питательными и пластическими веществами, а также энергией, необходимой для поддержания жизнедеятельности и продолжения рода, пищеварение выполняет важную барьерную и информационную функции. Первая проявляется в нарушении антигенных свойств потреблённых белков и избавлении организма от вредных метаболитов и токсинов, вторая связана с тем, что характер и свойства воспринятой пищи содержат для насекомых столь же существенные сведения о внешнем мире, как и звуки, формы, краски.
Отыскивая пищу, насекомые обычно руководствуются зрительными (визуальными) и обонятельными (ольфакторными) стимулами, воспринимая их по-разному в зависимости от насыщения или голода, физиологического состояния и ситуации. Непосредственно перед линькой или вскоре после неё, при заполнении зрелыми яйцами полости брюшка, насекомые обычно не реагируют на пищевые стимулы, отличаясь в любое другое время суточной ритмикой в остроте их восприятия или полным безразличием к ним при афагии (способность вообще не питаться на тех или иных фазах жизненного цикла). Немногие хищники, например личинки божьих коровок, воспринимают пищевые стимулы лишь при непосредственном контакте с потенциальной жертвой.
Причины пищевой специализации и степень её проявления различны. Некоторые хищные личинки мух не способны к быстрым движениям и довольствуются малоподвижными тлями. Нападение богомолов, напротив, провоцируется движениями жертвы в поле их зрения; они не способны питаться неподвижными червецами или щитовками, даже находясь среди них. Многие перепончатокрылые, парализующие свои жертвы молниеносным уколом яйцеклада, чаще всего бывают олигофагами и монофагами, так как от размера жертвы и свойственных ей способов самообороны зависит успех поединка, и лишь инстинктивная точность движений обеспечивает победу. Многие паразиты отыскивают хозяев, ориентируясь на свойственные последним места обитания и кормовые растения, но особенно интересна и сложна пищевая специализация фитофагов.
Обладая высокосовершенными органами чувств — рецепторами и руководствуясь разнородными стимулами в выборе кормового объекта, многие насекомые-фитофаги питаются на заведомо ядовитых растениях, которые издавна употребляются для производства инсектицидов типа пиретринов, ротенона и никотина. Не обнаруживая признаков отравления, табачный бражник Protoparce sexta L. весьма быстро выводит никотин из своего организма (за 30 минут — до 95 % воспринятого с пищей), а немногие из насекомых, воспринявших цианиды розоцветных, благодаря ферменту роданазе расщепляют и преобразуют их в безвредные тиоционаты.
Ядовитые глюкозиды горчичного масла даже привлекают бабочек-белянок (Pieridae) и капустную моль Plutella maculipennis Curt. В первом случае к такому глюкозиду относится синигрин, во втором — прогоитрин, но оба они не имеют никакой пищевой ценности. Предлагая гусеницам капустной белянки (Pieris brassicae L.) в изобилии любой корм, лишённый, однако, синитрина, мы обрекаем их на голодную смерть. Для колорадского жука привлекателен ядовитый соланин, но губителен близкий к нему демиссин. Короедов и заболонников влекут терпены древесных пород, запах которых для человека сходен с ароматом ландыша или сирени.
Установлено, что пищевыми стимулянтами для насекомых-фитофагов могут быть различные соединения, содержащиеся в растениях, но не представляющие собой ценности как пища. К их числу относятся разнообразные глюкозиды, терпеноиды, флавоноиды и многие другие привлекающие или, наоборот, отталкивающие насекомых вещества. Вместе с тем, охотно поглощая в больших количествах корм, богатый привлекающими соединениями (аттрактантами), насекомые быстрее развиваются и растут, и в этом смысле пищевую ценность таких соединений можно сравнить с действием пряностей и приправ.
Своеобразно отношение к питанию, выявленное у тутового шелкопряда (Bombyx mori L.). У данного вида к дистантному пищевому аттрактанту, позволяющему избирать пищу, относится цитраль из группы терпенов. Однако присутствия этого вещества недостаточно для отгрызания насекомым листа шелковицы, которое провоцируется контактными аттрактантами: аситостерином и флавоноидами. Заглатывание происходит только в том случае, если в состав листа входят кремниевая кислота и целлюлоза.
Близкие явления отмечены и у некоторых других насекомых, что предполагает определённую последовательность действия стимулов, обеспечивающих приём пищи. Если положительная или отрицательная ориентация на корм определяется присутствием соответственно аттрактантов и репеллентов, то начало питания или отказ от избранной пищи регулируется пищевыми инициантами и супрессантами. Для продолжения или прекращения питания необходимы стимулянты либо детерренты. Столь сложная регуляция приёма пищи зависит, в свою очередь, от степени насыщения насекомых, причём голод обостряет те из органов чувств, которые связаны с поиском и регистрацией пищи. Например, голодающие в течение 24 часов мухи в этом отношении активнее сытых примерно в 4 раза.
У мухи Phormia regina Mg. органы вкуса расположены на лапках и хоботке. Они весьма чувствительны к различным сахарам, но по мере насыщения их чувствительность понижается. Если давать мухам все более концентрированные растворы, то можно заставить их питаться снова и снова, вплоть до очередного снижения чувствительности и адаптации к стимулу. Однако повышение или снижение порогов чувствительности вообще не связывается с реальным насыщением. В частности, предлагая мухам хорошо усваиваемую ими глюкозу и совершенно неусваиваемую D-арабинозу, в обоих случаях можно наблюдать поведение, характерное для сытых особей, но во втором случае вероятна гибель мух от голода.
Наряду со вкусовыми рецепторами насыщение регулируют стимулы, исходящие от растягивающегося при питании брюшка. При нарушении нервных связей рецепторов растяжения мухи продолжают питаться вплоть до гибели от разрыва кишечника.
Таким образом, состояние голода и насыщения стимулируется влиянием вкусовых рецепторов лапок и хоботка и тормозится влиянием рецепторов растяжения кишечника.
Пищевые потребности.
В пищевой специализации насекомых выражаются их потребности в пище, которые определяются характером обмена веществ, набором ферментов в организме и способностью к самостоятельному синтезу некоторых важных органических соединений.
Наиболее очевидны потребности в белках, точнее, в слагающих их аминокислотах, которые выступают как строительный материал для тканей и клеток тела, ферментов, гормонов и других биологически активных соединений. Обычно они поступают с пищей; некоторые синтезируются в теле: незаменимые аминокислоты у насекомых, как и у позвоночных животных, — аргинин, валин, лейцин, изолейцин, триптофан, фенилаланин и др.
Менее выражена потребность в жирах, которые могут синтезироваться самим насекомым. Лишь линолевая и линоленовая кислоты, холестерин или другие стерины, необходимые для его синтеза, незаменимы. Выступая как структурные компоненты биологических мембран, резервный источник энергии и метаболической воды, они необходимы и для синтеза экдизона — гормона линьки насекомых.
Углеводы — важнейший источник энергии — либо воспринимаются с пищей, либо синтезируются из жиров и липидов. Многие хищники не нуждаются в поступлении углеводов извне, но фитофаги обычно получают их в избытке.
Не нуждаясь в нуклеиновых кислотах, синтезируемых всеми живыми организмами, некоторые насекомые усиливают процессы роста и развития при добавлении в диету пуринов и пиримидинов. Более определенна их зависимость от минеральных веществ (ионов K+, Ca2+, P3+, Mg2+, Zn2+ и др.), но в особенности от витаминов. Насекомые характеризуются явной и специфической потребностью в водорастворимых витаминах группы В, выступающих как коферменты. Не синтезируемые самим насекомым, они воспринимаются с пищей или от специфических кишечных симбионтов, способных к синтезу различных веществ. Особенно важны тиамин (B1), рибофлавин (В2), никотиновая кислота (РР), карнитин (Вt), инозит, тогда как отсутствие пиридоксина (В6) и фолиевой кислоты во многих случаях не приводит к заметным последствиям. Из витаминов других групп следует указать на заимствуемый из растений жирорастворимый витамин А и витамин С, которые некоторые насекомые синтезируют сами. Такие насекомые, как большой мучной хрущак (Tenebrio molitor L.) и ряд других вредителей запасов зерна, способны, питаясь сухим кормом, использовать метаболическую воду, образующуюся при окислении углеводов и жиров.
Слюнные железы.
Слюнные железы насекомых развиваются как парные эктодермальные органы в мандибулярном, максиллярном и лабиальном сегментах, но нередко их число сокращается до двух или одной пары либо они редуцируются. Наиболее постоянны и типичны гроздевидные лабиальные железы, от секреторных долек которых отходят тонкие протоки. Сливаясь в общие латеральные протоки, часто расширенные в объемистые резервуары, они выводят жидкую слюну в полость саливария через отверстие у основания гипофаринкса (рис. 79).
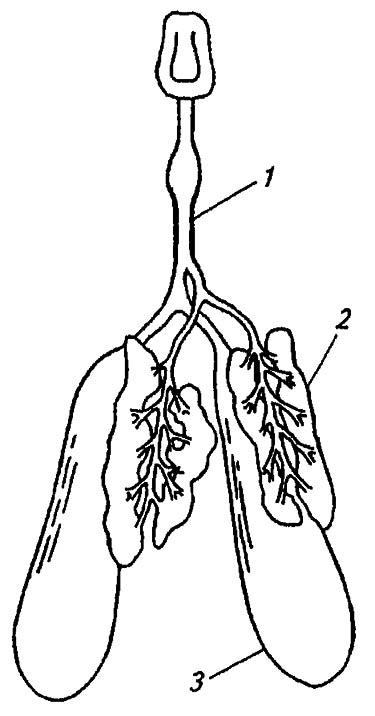
Рисунок 79. Слюнные железы насекомых (по Romoser, 1981):
1 — проток; 2 — лабиальные железы; 3 — резервуар
Обычно в слюне содержатся ферменты, расщепляющие углеводы, — карбогидразы (амилаза, инвертаза), а у некоторых хищников — липазы и протеиназы, переваривающие соответственно жиры и белки. В слюне тлей находится пектиназа, разрушающая пектиновые оболочки клеток и облегчающая проникновение стилетов в ткани растений, в слюне некоторых клопов — шалуронидаза, действующая сходным образом на соединительную ткань. Для кровососов характерно присутствие в слюне антикоагулянтов, препятствующих свёртыванию крови, агглютинов, действующих на её клетки, а также анестезирующих веществ. Иногда слюна ядовита или содержит вещества типа ауксинов. Содействуя образованию галлов — опухолевидных разрастаний ткани растения вокруг питающегося насекомого, — эти вещества обильно обеспечивают его пищей и надёжной дополнительной защитой.
Слюнные железы нередко преобразуются и начинают продуцировать феромоны, шёлк или аллотрофические вещества, предназначенные для питания других особей. Например, мандибулярные железы пчелиной матки выделяют вещества, тормозящие развитие яичников всех прочих самок, а железы рабочих особей — маточное молочко. Своеобразны аллотрофические слюнные железы самцов скорпионницы Panopra, заполняющие тремя парами толстых трубок почти все тело. Наряду с обильной бурой слюной, извергаемой при питании и при обороне, они способны образовывать молочно-белые пакетики, которые самка охотно поедает при спаривании.
Особенно примечательны шёлкоотделительные железы гусениц тутового шелкопряда. Образованные извитыми секретирующими трубочками, они продолжаются в расширенные резервуары и далее в более тонкие протоки, впадающие с обеих сторон тела в волочильню, или прядильный пресс (рис. 80). Секретируемый этими железами белок фиброин составляет основу шелковинки, которая, переходя в резервуар, облекается слоем серицина. Выдавливаемая из пресса через прядильный сосочек на нижней губе гусеницы шёлковая нить используется для постройки кокона.

Рисунок 80. Шелкоотделительные железы тутового шелкопряда (по Шванвичу, 1949)
Растворимый в горячей воде серицин удаляется при промышленной обработке коконов шелкопряда; это облегчает их разматывание в нить, достигающую 1 500 м в длину. Готовое шёлковое волокно не уступает по прочности стали и, выдерживая нагрузку на разрыв до 50 кг на 1 мм2 поперечного сечения, значительно превосходит по прочности шерстяную и хлопковую нити.
Кишечный тракт.
Откусанная и смятая жвалами или всосанная через хоботок пища вместе со слюной поступает в цибарий и далее в глотку. Глотка имеет мускулистые стенки, а у некоторых насекомых преобразована в глоточный насос; она переводит пищевой комок в кишечный тракт, схема которого изображена на рисунке 81.

Рисунок 81. Схема кишечного тракта насекомых (по Шванвичу, 1949):
1 — слюнные железы; 2 — глотка; 3 — пищевод; 4 — зоб; 5 — провентрикулус; 6 — кардиальный клапан; 7 — перитрофическая мембрана; 8 — мальпигиев сосуд; 9, 10 — соответственно пилорический и ректальный клапаны; 11 — анус
Обычно кишечник разделён на три отдела: переднюю, среднюю и заднюю кишку, которые, различаясь по происхождению, обособлены друг от друга клапанами. Переднюю кишку отделяет от средней кардиальный клапан, среднюю от задней — пилорический. Передняя и задняя кишка образуются из эктодермы и, как многие другие её производные, имеют тонкую кутикулярную выстилку. Энтодермальная средняя кишка лишена кутикулы.
Передняя кишка.
Вслед за глоткой, находящейся в начале передней кишки, расположен тонкий пищевод, нередко переходящий без видимых границ в расширенный зоб. У многих насекомых пищевод имеет дивертикулы — отходящие от его стенки выросты, и пищевой резервуар — объёмистый мешок, соединённый с пищеводом узкой трубочкой. Эти образования так же, как и зоб, обычно служат для накопления и резервирования запасов пищи, перевариваемой здесь лишь ферментами слюны. Например, у медоносных пчёл цветочный нектар насыщается в зобу ферментами, под воздействием которых он преобразуется в мед, а у хищных жужелиц и прямокрылых проникающие сюда ферменты средней кишки способны переваривать белки. Пищеварительная функция зоба особенно полно выражена у форм с внекишечным пищеварением, вводящих протеолитические ферменты со слюной в тело пойманной, но ещё не проглоченной жертвы. Однако всасывания продуктов переваривания пищи здесь не происходит, и основным назначением пищевода и зоба служит проведение пищевого комка.
Если у насекомого имеется развитый пищевой резервуар, в него обычно поступают различные жидкости, а твёрдая пища переходит в зоб и далее в провентрикулус. Самки кровососущих комаров, например, прокалывая стилетами кожу, направляют всосанную кровь в зоб и провентрикулус, а потребляя воду, заполняют ею резервуар. Акт прокола определяет путь следования пищи, и если самке предложить свободную капельку свежей крови, то она погибнет после свёртывания крови в пищевом резервуаре.
Наиболее деятельный компонент передней кишки — провентрикулус, иногда называемый мышечным желудком (рис. 82). Сильно развитые кольцевые мышцы и кутикулярные зубцы, вдающиеся в его полость, обеспечивают эффективное перетирание и фильтрацию пищевых частиц, дополняя тем самым функции челюстей. Иногда движения провентрикулуса, видимые через прозрачные покровы некоторых личинок, напоминают движения глотки: вдаваясь в пищевой комок, находящийся в зобу, он как бы «откусывает» некоторую часть этого комка и приступает к методичному «пережёвыванию». Отдельные порции «пережёванной» пищи провентрикулус передаёт через кардиальный клапан в среднюю кишку.
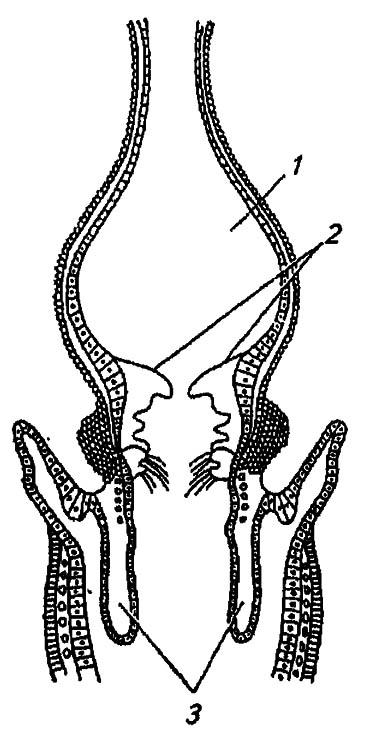
Рисунок 82. Схема строения провентрикулуса насекомых (по Romoser, 1981):
1 — зоб; 2 — кутикулярные зубцы; 3 — кардиальный клапан
Кардиальный клапан кольцевидной складкой вдаётся в полость средней кишки. При этом, начиная с места перегиба складки, кутикулярная выстилка исчезает, и, очевидно, именно здесь проходит граница передней и средней кишки. Полагают, что клапан препятствует обратному ходу пищи, но у многих насекомых содержимое средней кишки все же проникает в переднюю. У некоторых насекомых кардиальный клапан участвует в выделении перитрофической мембраны, облекающей тончайшим чехлом пищевые массы и защищающей от повреждения нежный эпителий средней кишки.
Перитрофическая мембрана.
Она образована сетью микрофибрилл и тончайшей белково-хитиновой плёнкой в её ячейках. Легко проницаемая для воды и растворов минеральных солей, она обладает свойствами одностороннего фильтра в отношении более крупных молекул. В частности, молекулы пищеварительных ферментов легко проходят к перевариваемой ими пище, но не способны вернуться обратно. Лишь продукты полного расщепления достигают эпителия средней кишки и абсорбируются его клетками. Даже некоторые дисахариды и простые липиды, как и многие микроорганизмы, не способны преодолеть барьер перитрофической мембраны.
Весьма обычная для насекомых перитрофическая мембрана отсутствует у форм, отличающихся внекишечным перевариванием пищи и предельно суженным рационом легкоперевариваемых веществ. Там же, где она есть, её структура однообразна, хотя по способу образования выделяют несколько типов мембран, в частности обычный и зональный. В первом случае перитрофическая мембрана выделяется всей поверхностью эпителия средней кишки и как бы отслаивается от него, во втором — узкой зоной специализированных клеток кардиального клапана (рис. 83), как, например, у личинок двукрылых, некоторых взрослых мух и, вероятно, уховёрток и термитов.
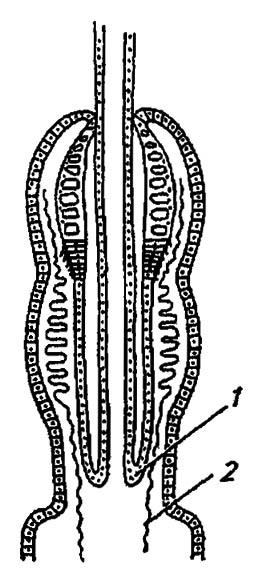
Рисунок 83. Кардиальный клапан (1) и образование перитрофической мембраны (2) (по Шванвичу, 1949)
Средняя кишка.
Облеченная перитрофической мембраной, основательно «пережёванная» и отфильтрованная пищевая масса поступает в среднюю кишку. Здесь она подвергается воздействию ферментов и расщепляется на относительно простые органические соединения, либо абсорбирующиеся клетками эпителия, либо диффундирующие в гемолимфу непосредственно через стенку кишечника. Далее они включаются в общий цикл биохимических превращений, происходящих в жировом теле, гемолимфе и других тканях тела насекомого. Таким образом, средняя кишка — основной орган переваривания пищи и всасывания образовавшихся при этом продуктов.
В исходном состоянии средняя кишка имеет вид прямой трубки, снабжённой близ её переднего и заднего края слепыми выростами для увеличения активной поверхности всасывания. Однако у подавляющего большинства насекомых она удлиняется, образуя изгибы и петли, а у высших двукрылых свёртывается плотной двойной спиралью (рис. 84).
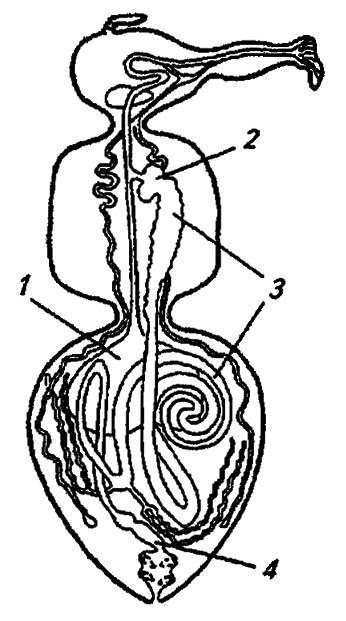
Рисунок 84. Кишечный тракт высших двукрылых (по Шванвичу, 1949):
1 — пищевой резервуар; 2 — провентрикулус; 3, 4 — соответственно средняя и задняя кишка
Как правило, средняя кишка особенно велика у тех форм, которые питаются плохо усвояемыми субстратами, например у копрофагов, тогда как у плотоядных она относительно коротка. У клопов и некоторых других насекомых средняя кишка дифференцирована на зоны, отличающиеся по форме и строению эпителия (рис. 85).
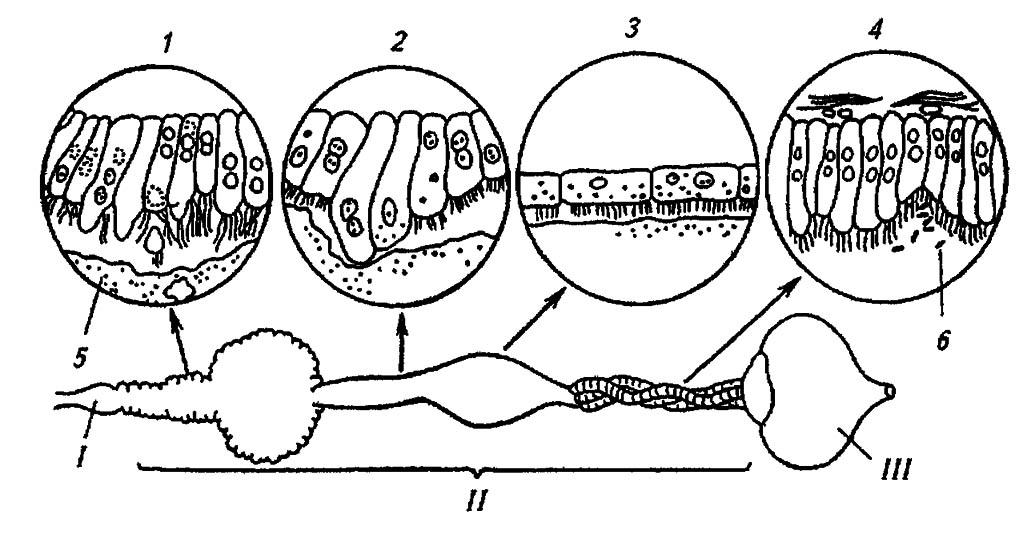
Рисунок 85. Дифференциация зон средней кишки у вредной черепашки. В кружках — последовательные зоны (1–4) средней кишки при большом увеличении (по Тьпценко, 1976):
I, II, III — соответственно передняя, средняя и задняя кишка; 5 — перитрофическая мембрана; 6 — симбионты
Эпителий средней кишки (рис. 86) обычно представлен столбчатыми клетками, на активной поверхности которых находится рабдориум — слой неподвижных цитоплазматических палочек, подостланный складчатой базальной мембраной. Как и все секретирующие клетки, они особенно богаты эндоплазматическим ретикулумом, рибосомами и митохондриями. Среди клеток эпителия рассеяны регенеративные гнезда — крипты. Сосредоточенные в них мелкие, интенсивно делящиеся клетки содействуют обновлению эпителия и восстанавливают его при линьках и метаморфозе насекомых. У многих личинок эпителий средней кишки сбрасывается в просвет кишечника и замещается провизорным (временным) эпителием куколок, который, в свою очередь, уступает место дефинитивному (постоянному) имагинальному эпителию. Однако при глубоком метаморфозе у жуков, бабочек и мух куколочный эпителий вообще не формируется.
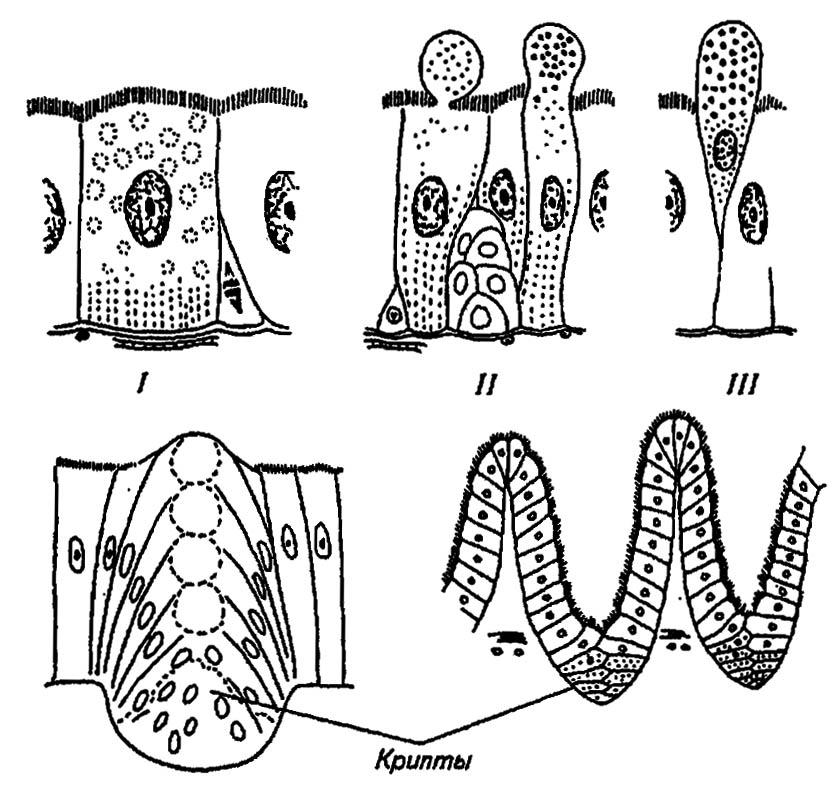
Рисунок 86. Эпителий средней кишки и различные типы секреции (по Тыщенко, 1976):
I, II, III — соответственно апокринный, мерокринный, голокринный типы секреции
Назначение эпителиальных клеток средней кишки — секреция ферментов и всасывание продуктов гидролиза и распада белков, углеводов и жиров. Секреция ферментов может происходить без видимых изменений клеток за счёт выделения накопленных в них ферментов между палочками рабдориума. Этот так называемый апокринный тип секреции нередко сочетается с мерокринным и голокринным типами. В первом случае содержащие ферменты вакуоли вместе с частью цитоплазмы отшнуровываются от клеток и затем изливаются в полость кишечника, во втором — заполненные секретом клетки целиком выходят из эпителия и, погибая, освобождают содержимое своих тел.
У вредной черепашки (Eurygaster integriceps Put.) секреция происходит в первой зоне средней кишки (см. рис. 85) и частично во второй и третьей зонах, тогда как клетки четвёртой зоны вмещают симбиотические микроорганизмы. У мухи цеце (Glossinapalpalis L.), передающей человеку опаснейшую сонную болезнь, все процессы секреции сосредоточены в средних зонах.
Всасывание продуктов распада происходит либо повсеместно, либо на отдельных участках средней кишки. Например, у колорадского жука всасывание углеводов сосредоточено в передней половине средней кишки, а продуктов расщепления жиров — только в задней половине. У других насекомых жиры абсорбируются в дивертикулах, но у пустынной саранчи (Schistocerca gregaria Forsk.) здесь всасываются только аминокислоты. Нередко в первой половине средней кишки насекомых всасывается вода, которая затем выделяется в задней половине, создавая тем самым необходимый для пассивного транспорта молекул углеводов и липидов градиент концентрации. Всасывание аминокислот требует активного транспорта и часто происходит вопреки градиенту концентрации.
Выделение ферментов и всасывание продуктов пищеварения регулируются в зависимости от физиологического состояния организма.
Отмечено, что сам акт питания и заполнение пищей зоба стимулируют выделение секретов и ферментативную активность; все эти процессы контролируются также импульсами стоматогастрической нервной системы, регистрирующей растяжение брюшка, и, видимо, гормонами головного мозга. Переваривание пищи требует совершенно определённой реакции среды кишечника (рН 6–8) и перистальтики, которая, в свою очередь, регулируется либо посредством нервной системы и гормонов, либо за счёт возбуждения рецепторов в окружающей кишечник мускулатуре, то есть миогенным путем.
Задняя кишка.
Она обособлена от средней пилорическим клапаном и обычно разделена на тонкую и прямую кишку с ректальным клапаном между ними (см. рис. 81). В ней имеются тонкая кутикулярная выстилка, проницаемая для воды, минеральных солей и некоторых органических соединений, а также мышечные волокна, расположение которых необычно: толстые кольцевые мышцы находятся под более тонкими продольными.
Рисунок 81. Схема кишечного тракта насекомых (по Шванвичу, 1949):
1 — слюнные железы; 2 — глотка; 3 — пищевод; 4 — зоб; 5 — провентрикулус; 6 — кардиальный клапан; 7 — перитрофическая мембрана; 8 — мальпигиев сосуд; 9, 10 — соответственно пилорический и ректальный клапаны; 11 — анус
Непосредственно за пилорическим клапаном расположен сфинктер, а в промежуток между ними впадают мальпигиевы сосуды. Сфинктер регулирует поступление непереваренных остатков пищи в заднюю кишку.
Важнейшие функции задней кишки — всасывание воды и формирование обезвоженных экскрементов, что имеет особое значение для поддержания водного баланса в условиях постоянного дефицита влажности. У многих сапрофагов и у питающихся древесиной ксилофагов тонкая кишка преобразуется в ферментативную камеру, где обитают разнообразные симбионты: сапрофитные бактерии, грибы и простейшие разных групп. Выделяя ферменты, расщепляющие целлюлозу до глюкозы, или продолжая разлагать не переваренные в средней кишке остатки пищи, они синтезируют некоторые незаменимые аминокислоты, витамины, стеролы и, по некоторым данным, способны к фиксации атмосферного азота как почвенные нитрификаторы. Лишённый симбионтов термит после дефаунации его кишечника кислородом вскоре гибнет от голода, хотя в нормальных условиях способен длительно существовать, питаясь, например, ватой, то есть практически чистой клетчаткой, которую разлагают его симбионты. Многие другие насекомые более легко расстаются со своими симбионтами, а щетинохвостки Ctenolepisma lineata F. переваривают клетчатку без их участия — собственными ферментами.
Выстланная кубическим эпителием, задняя кишка не выделяет собственных пищеварительных ферментов, но обычно снабжена 6–8 ректальными железами, всасывающими воду из экскрементов. Особой сложности эти железы достигают у двукрылых, причём для них характерна ритмическая пульсация. У некоторых червецов слепой вырост прямой кишки (ректальная ампула), образуя широкую петлю, срастается с передней кишкой в «фильтрационную камеру». У других равнокрылых эти камеры (рис. 87) формируются при срастании обоих концов средней кишки или при врастании мальпигиевых сосудов в область кардиального клапана.
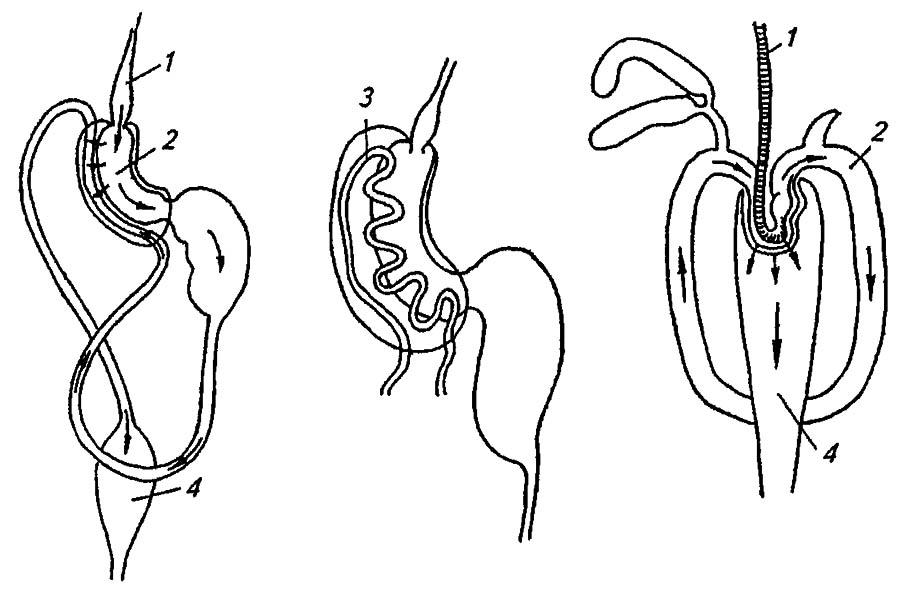
Рисунок 87. Фильтрационные камеры равнокрылых (по Шванвичу, 1949):
1 — пищевод; 2 — средняя кишка; 3 — мальпигиевы сосуды; 4 — задняя кишка
Таким образом, вода и балластные углеводы, обильно поступающие в кишечник при высасывании соков растений, отфильтровываются в самом его начале и в среднюю кишку поступает лишь относительно концентрированный и обогащённый белками раствор. Побочное следствие такого способа питания — выделение жидких сахаристых экскрементов, именуемых падью. Однако экскременты большинства насекомых представляют собой совершенно сухие пакетики, облечённые остатками перитрофической мембраны.
Длительность пребывания пищи в кишечнике различных насекомых варьирует в весьма широких пределах. Например, у таракана Periplaneta americana L. пищеварение продолжается около 20 ч, у близкого вида P. australasiae L. — 7, у рыжего таракана Blatella germanica L. — только 3 ч.
Таким образом, функции задней кишки насекомых многообразны. Наряду со всасыванием воды и формированием экскрементов здесь происходят ферментация и всасывание веществ, синтезируемых симбиотическими микроорганизмами, или эти вещества переводятся антиперистальтическими движениями кишечника в полость средней кишки. Всасывание воды, ионов и относительно простых органических соединений обычно происходит здесь вопреки градиенту концентрации путём активного транспорта молекул через мелкопористую кутикулу и эпителий. У зимующих насекомых и многих паразитических перепончатокрылых задняя кишка накапливает экскременты и выводит их наружу лишь с началом активного существования.

