Рецепция и поведение насекомых
Рецептор — элементарный орган чувств, образованный сенсорными нейроном или нейронами и вспомогательными структурами. Настроенный на восприятия адекватных раздражений, он обеспечивает взаимодействие их носителей со специфическими белковыми молекулами на плазматической мембране нейрона. В результате этих взаимодействий её проницаемость меняется и возникающие ионные потоки приводят к деполяризации мембраны, регистрируемой как постоянно нарастающий рецепторный потенциал. Последний содействует образованию импульсов возбуждения и их распространению в соответствующие нервные центры. Таким образом, рецептор трансформирует энергию раздражающего стимула в нервные импульсы, частоты которых соответствуют силе раздражения.
Малоклеточные и миниатюрные рецепторы насекомых отличаются исключительной чувствительностью и устойчивостью к помехам, надёжностью и экономичностью работы. По многим параметрам они превосходят измерительные приборы высокой точности. Так, одни из них, воспринимая летучие соединения в исчезающе малых концентрациях, служат «счётчиками» отдельных молекул, а другие регистрируют единичные фотоны или колебания субстрата с амплитудой, равной радиусу атома водорода.
Для насекомых характерны первичные рецепторы, нейроны которых не только воспринимают энергию стимула, но и сами генерируют и передают нервные импульсы. В этом отношении они противопоставляются вторичным рецепторам позвоночных животных, образованным двумя нейронами: воспринимающим раздражение и передающим возникающее в нем возбуждение через синаптическую щель на нейрон, генерирующий импульсы в центральную нервную систему.
После проведённой первичной обработки, кодирования и передачи информации её последующий анализ, а затем и синтез целостного образа раздражителя происходят в нервных центрах, которые вместе с органами чувств и проводящими путями объединяются в анализаторы.
У насекомых выявлено не менее девяти анализаторов: зрительный, обонятельный, вкусовой, слуховой, тактильный, двигательный, гравитационный, висцеральный и температурный. Имеются сведения о способности насекомых регистрировать изменения влажности и электромагнитного поля, но соответствующие нервные центры ещё не исследованы. Между тем перечисленному набору воспринимаемых ощущений отвечают лишь четыре класса рецепторов: механо-, термо-, хемо- и фоторецепторы. Объяснение этому несоответствию можно найти в том, что, например, различия вкусовых и обонятельных стимулов определяются лишь той средой — жидкой или газообразной, в которой воспринимаются их носители, а противопоставления тактильных, двигательных, гравитационных ощущений или ощущений звуковых волн и состояния внутренних органов (висцеральное чувство) сводятся к различиям в механических раздражениях сенсорного нейрона.
Механорецепция
Описание механорецепторов уместно начать с простейшего из них — осязательного волоска, именуемого трихоидной сенсиллой (рис. 119). Расположенный на покровах, в сочленовной резилиновой мембране, он связан с дендритом сенсорного нейрона, окончание которого (преобразованная ресничка) одето кутикулярным чехликом — сколопсом. Этот волосок настолько упруг, что любое его отклонение сразу передаётся мембране и, деформируя сколопс, генерирует рецепторный потенциал. Обычно сенсиллы такого типа группируются в местах наиболее частых контактов с механическими раздражителями и дифференцируются как органы осязания и ветрочувствительные волоски. Последние, как правило, более тонкие и подвижные.

Рисунок 119. Трихоидная сенсилла и её производные (по Шванвичу, 1949):
1 — сенсилла; 2 — нервы; 3 — колоколовидная сенсилла
При стимуляции трихоидной сенсиллы постепенно нарастающий рецепторный потенциал достигает порогового значения и возбуждает незатухающие нервные импульсы. Их частоты определяются силой воздействия на рецептор. По восприятию воздействия тоническим сенсиллам противопоставляются фазовые. При восприятии фазовыми сенсиллами высокие значения рецепторного потенциала и частот отмечаются только в начале воздействия и при длительном раздражении подавляются, при восприятии тоническими — сохраняются неизменными все время, пока действует раздражитель. Наряду с ними выделяют фазотонические сенсиллы, реагирующие в начале раздражения всплеском нервных импульсов, частоты которых, однако, резко снижаются при длительном действии раздражителя. Таким образом, тонические сенсиллы передают в нервные центры анализатора информацию о силе и длительности раздражения, фазовые — о его начале, а фазотонические объединяют свойства тех и других.
Обычно сенсиллы специализируются на определённом режиме восприятия стимула, но в яйцекладе некоторых мух присутствуют все три их типа. Видимо, это позволяет им отличать кратковременные и случайные соприкосновения с субстратом при движении от более длительных контактов.
Особенно густые скопления фазотонических трихоидных сенсилл (волосковые пластинки) в сочленениях и на границах сегментов регистрируют взаимные смещения и положения последних. Являясь проприоцепторами (рецепторами собственного положения), эти сенсиллы нередко приобретают значение рецепторов гравитации и, воспринимая ориентацию тела в гравитационном поле, содействуют коррекции его положения при полете и плавании.
Несмотря на простоту строения, трихоидные сенсиллы участвуют в рецепции весьма разнообразных механических стимулов. В этом отношении они противопоставляются кампаниформным (колоколовидным) сенсиллам, специализирующимся как проприоцепторы упругих деформаций кутикулы при мышечных сокращениях. Морфологически они отличаются от трихоидных сенсилл только тем, что лишены волоска, преобразованного здесь в невысокий овальный колпачок. Располагаясь по линиям деформаций, они передают в фазотоническом режиме сигналы о силе и направленности изгибаний кутикулы.
В еще большей степени специализированы хордотональные сенсиллы — сколопидии, которые наряду с функциями проприоцепто-ров нередко приобретают способность к регистрации звуков. Дендрит сенсорного нейрона сколопидия образует тонкую, расширенную посредине ресничку. Окруженная кутикулярным сколопсом, выделяемым обкладочной клеткой, она простирается за его пределы в центральный канал колоколовидной шапочки (рис. 120), к которой примыкает шапочковая клетка. Многочисленные тонофибриллы этой клетки содействуют прикреплению всего комплекса к эпикутикуле покровов и при их растяжении переводят раздражение на ресничку дендрита. Обычно сколопидии группируются в более или менее сложные хордотональные органы. Натянутые, как струны, они регистрируют малейшие деформации и нередко дифференцируются на относительно мелкие тонические и более крупные фазовые и фазотонические элементы.

Рисунок 120. Хордотональная сенсилла насекомых (по Gillot, 1980):
1 — сенсорный нейрон; 2 — сколопс; 3 — кутикула; 4, 5 — соответственно шапочковая и обкладочная клетка
О конкретном назначении механорецепторов можно судить по следующим примерам. Богомол Mantis religiosa L., заметив жертву, остаётся неподвижным и лишь слегка поворачивает вслед ей голову. Раздражая при этом волосковые пластинки в шейном сочленении, он легко перестраивает направление броска. Однако выключение этих рецепторов, не препятствуя слежению, сокращает точность бросков (прицельность) в несколько раз. Ещё более нагляден тарзальный рефлекс, свойственный многим насекомым. Он заключается в следующем. Отрыв лапок насекомого от опоры сразу вызывает полет, который прекращается столь же быстро при возобновлении контакта с субстратом.
Вместе с тем для поддержания полёта необходима стимуляция ветрочувствительных сенсилл на голове и крыльях, а для его контроля — проприоцепторов антенн: чем сильнее на них давление встречных потоков воздуха, тем меньше амплитуда взмахов крыльев. Не менее важную роль играют механорецепторы в поддержании равновесия и регистрации положения тела в поле гравитации. Сопоставляя раздражения от разных частей тела, соответствующие анализаторы весьма тонко отличают малейшие (до 2°) отклонения от горизонтали. При этом у водных форм роль проводников гравитационных стимулов нередко исполняют пузырьки воздуха. Стремясь к поверхности и оказывая давление на расположенные над ними группы сенсилл, они содействуют также сохранению равновесия и регистрации глубины погружения под воду.
Все эти столь разнообразные функции осуществляются осязательными волосками и развивающимися на их основе более сложными кампаниформными и хордотональными сенсиллами. Дальнейшая специализация последних как виброрецепторов приводит к оформлению весьма чувствительных к малейшим сотрясениям субстрата подколенных органов кузнечиков и расположенных в антеннах джонстоновых органов. Выполняя роль проприоцепторов, они одновременно приобретают способность к восприятию звуков. На этом основании их можно противопоставить всем ранее рассмотренным контактным механорецепторам, воспринимающим механические стимулы лишь при непосредственном контакте с их носителями как рецепторы дистантного действия, работающие на расстоянии от источника стимула. Поэтому мы рассмотрим их вместе со специализированными органами слуха, а сейчас остановимся на рецепторах растяжения.
В отличие от прочих биполярных сенсорных нейронов мультиполярные нейроны рецепторов растяжения распластываются своими многочисленными дендритами по поверхности внутренних органов и в полости тела. Регистрируя натяжение мышц, они генерируют импульсы, частота которых возрастает при усилении и ускорении растяжения. При этом в соответствующие анализаторы передаётся информация о силе и скорости сокращения мышц. Наряду с регистрацией состояния мускулатуры рецепторы такого рода сообщают в нервные центры о перистальтике кишечника, биении спинного сосуда, движениях внутренних органов и активности вентиляции трахей.
Восприятие и генерация звуков
Способность к восприятию и генерации звуковых волн у разных видов насекомых далеко не одинакова. Не ограничиваясь восприятием сигналов, насекомые отвечают своему партнёру, но гораздо чаще безмолвно реагируют на них специфическими актами поведения.
О биологическом значении акустической коммуникации можно судить по набору выявленных сигналов, среди которых различаются призывы, обращеёные к самкам, и их ответы, брачные песни спаривающихся особей, звуковые сигналы самцов, охраняющих избранную территорию, и сигналы агрессии, предшествующие поединкам, сигналы протеста, бедствия и тревоги.
Наибольшую акустическую одарённость обнаруживают прямокрылые, в особенности самцы сверчков, которые превосходят в этом отношении не только рыб, амфибий и рептилий, но и многих певчих птиц. Общее число сигналов, связанных со специфическими актами поведения, достигает у насекомых 13 (у отдельных видов — 7).
Насекомые владеют различными средствами и способами производства звуков. Весьма обычны для них фрикционные органы, принцип работы которых основан на трении острого кушкулярного канта о зазубренную поверхность. Например, у кузнечиков и сверчков соответствующие структуры расположены на передних крыльях: потирая их друг о друга, они издают характерное стрекотание (стридуляцию). Сходный эффект достигается трением о край крыла внутренней поверхности бёдер задних ног, которая у саранчовых несёт мелкие шипики.
Свойственные певчим цикадам тимбальные органы имеют жёсткие кутикулярные мембраны, которые приводятся в действие сильными асинхронными мышцами. Вгибаясь и выгибаясь, как дно у жестяной банки, они генерируют звуки, многократно усиливаемые резонатором, функции которого выполняет брюшко, пустое у поющих самцов.
Звуки, возникающие при вибрации крыльев и стенок грудных сегментов, — побочное следствие работы крыловых мышц, но для многих двукрылых эти звуки приобретают значение видоспецифичных сигналов и акустических ориентиров.
Засасывая воздух в кишечник и с силой выталкивая его наружу, бражник мёртвая голова (Acherontia atropos L.) шипит, как потревоженная змея. Некоторые таракановые, перепончатокрылые и двукрылые шипят, выпуская воздух через дыхальца. Многие жуки-точильщики призывают самок стуком головы о стенки прокладываемых в мёртвой древесине ходов. Эти звуки, напоминающие тихое тиканье часов («часы смерти»), нередки в старых деревянных строениях. Сходным образом предупреждают друг друга об опасности термиты. Голодные личинки некоторых ос, требуя пищи, скрежещут челюстями о края ячеек гнезда. Многие совки издают ультразвуки, узнавая приближающуюся летучую мышь с расстояния до 30 м или воспринимают её собственные ультразвуковые посылки с ещё больших расстояний.
Звуки, производимые насекомыми, варьируют по частоте от 5 до 2 кГц, а их сила у наиболее громкоголосых кузнечиков и цикад достигает 110–115 дБ. Нередко они слагаются в песни, образованные элементарными звуковыми посылками — пульсами и щелчками, длящимися обычно 10–30 мс, сериями пульсов и непрерывными трелями. При этом наборы пульсов в сериях и трелях варьируют от 5 до 800 за 1 с.
Частотные характеристики генерируемых звуков в основном определяются механическими свойствами звукопроизводящих органов, тогда как временной рисунок песен контролируется работой нервно-мышечного аппарата и центральной нервной системой. Однако прежде чем судить о значении тех или иных параметров звуковых сигналов для акустической коммуникации, необходимо ознакомиться со свойствами органов слуха насекомых. При этом заметим, что способность звуковых волн распространяться в воздухе, а также в водной и твёрдой средах не позволяет провести резких границ между слухом и виброрецепцией. Очевидно, речь может идти лишь о противопоставлении дистантной и контактной рецепции механических стимулов.
Один из наиболее сложных хордотональных органов, способных к восприятию звуковых волн в воздушной и водной средах, — джонстонов орган. Обычно он образован группой из 1—3-нейронных сколопидиев, которые прикрепляются к сочленовной мембране 2-го и 3-го члеников антенн. Воспринимая колебания жгута, они передают импульсы возбуждения непосредственно в головной мозг. У мух-дрозофил этот орган настроен на видоспецифичные сигналы, порождаемые вибрацией крыльев самцов, а у жуков-вертячек наряду с функциями виброрецептора он регистрирует искривления поверхностной плёнки воды (мениски) на расстоянии до 15 мм от преграды.
Особенно сложен и своеобразен джонстонов орган у комаров (рис. 121). Его основу составляют тысячи нейронов, которые группируются концентрическими слоями у основания жгута антенн. Воспринимая малейшие вибрации и отклонения антенн, они генерируют импульсы в фазовом и тоническом режимах. Если при этом фронт звуковых волн параллелен оси антенны, то покрывающие её волоски начинают вибрировать и, раскачивая жгут антенны, вызывают максимальное раздражение рецепторов. Воспроизводя посредством звуковых генераторов специфические для комаров сигналы, в некоторых случаях удаётся привлечь этих докучливых насекомых к засасывающим ловушкам.

Рисунок 121. Джонстонов орган комара (по Жантиеву, 1977):
1 — нервы; 2 — жгут антенны; 3 — джонстонов орган; 4 — второй членик антенны
Специализированные органы слуха насекомых снабжены тимпанальными мембранами, регистрирующими звуковое давление и его градиент. Отмеченные у прямокрылых, цикад, некоторых клопов, сетчатокрылых и бабочек, они различаются положением, набором рецепторов и вспомогательными структурами. У кузнечиков, сверчков и медведок тимпанальные органы расположены на голенях передних ног и открываются наружу узкими окошечками. Основу этих органов составляют разграниченные стволами трахей две тимпанальные мембраны и примыкающие к ним группы сколопидиев подколенного органа, промежуточного отдела и слухового гребня (рис. 122).
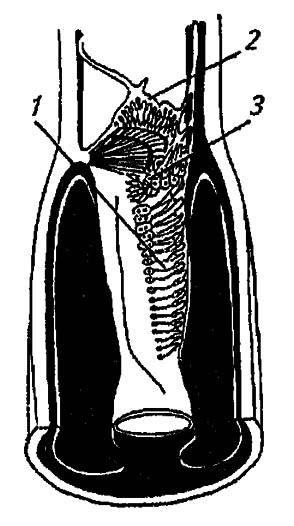
Рисунок 122. Слуховой гребень кузнечиков (по Шванвичу, 1949):
1 — слуховой гребень; 2 — подколенный орган; 3 — промежуточный отдел
Подколенный орган, образованный веером сколопидиев, служит высокочувствительным виброрецептором. Немногочисленные сенсиллы промежуточного отдела, натянутые между кутикулой и трахеями, вероятно, выполняют функции проприоцепторов. Расположенные в один ряд сколопидии слухового гребня своеобразны в двух отношениях: во-первых, они примыкают к трахее и не скрепляются шапочковыми клетками с кутикулой; во-вторых, постепенно уменьшаются в размерах, начиная с самых крупных проксимальных и кончая мелкими дистальными. Не имея контакта с тимпанальной мембраной, они воспринимают колебания, передаваемые ею на трахейные стволы.
У саранчовых дендриты сенсорных нейронов вплотную прилегают к мембране и возбуждаются ее колебаниями непосредственно. В отличие от органов кузнечиков тимпанальные органы саранчовых, а также бабочек и цикад расположены в первом сегменте брюшка или в заднегруди. У некоторых бражников органами слуха служат заполненные трахейными мешками членики нижнегубного щупика, соприкасающиеся с рудиментом верхней губы — пилифером, а у сетчатокрылых и бабочек-бархатниц — заполненные гемолимфой вздутия при основаниях радиальных жилок крыла.
Обычно в тимпанальные органы насекомых входят десятки, реже — сотни нейронов, но у цикад их число достигает 1 500. Реагируя на раздражения, они генерируют залпы импульсов, частоты которых вскоре сокращаются и стабилизируются на некотором уровне. Обладая сравнительно невысокой остротой слуха (7–40 дБ), они вместе с тем способны различать звуковые посылки, следующие друг за другом с частотой 100–300 в секунду. Несмотря на рекордное число нейронов, тимпанальные органы цикад настроены лишь на определённый диапазон частот, соответствующий максимальной энергии в спектре призывных песен самцов. У саранчовых выявлено четыре типа разных нейронов, настроенных на восприятие низких и высоких частот. У способных генерировать и воспринимать ультразвук бабочек-совок выявлено три нейрона, причём только два из них возбуждаются ультразвуком, а третий, наоборот, тормозит импульсную активность. Вероятно, это торможение повышает «внимательность» всего органа к восприятию ультразвуковых посылок летучих мышей.
Таким образом, тимпанальные органы насекомых воспринимают в первую очередь не частотные, а временные параметры звуковых стимулов, передавая в высшие нервные центры информацию о длительности импульсов и о характере их повторений, то есть временной рисунок песен.
Преобразованная сенсорными нейронами информация о характере звуковых стимулов поступает в нервные центры грудных ганглиев, где подвергается дальнейшей обработке. Расположенные здесь ассоциативные нейроны, работая в разных режимах, отвечают на одни и те же сигналы по-разному. Фазовые нейроны регистрируют начало песен и на каждый сигнал генерируют по 1–2 импульса, подчёркивая ритмическую организованность звуковых посылок. Одновременно тонические нейроны регистрируют длительность отдельных сигналов.
Таким образом, интенсивность импульсов, продуцируемых сенсорными нейронами в зависимости от интенсивности раздражителя, определяет частоту импульсации ассоциативных нейронов и вместе с тем общее число реагирующих элементов в нервном центре. Некоторые нейроны при повышении интенсивности опосредованных сигналов тормозятся и выключаются, другие — вступают в обработку акустической информации. У саранчи Locusta migratoria L. соответствующие нейроны могут работать либо в фазовом, либо в тоническом режиме. При звуках средней и низкой интенсивности они дают фазовые ответы, при звуках высокой интенсивности — тонические. Это позволяет различать ритмические посылки и обусловливает высокую чувствительность анализатора к изменениям интенсивности шумовых «фоновых» звуков.
Очевидно, что на уровне нервных ганглиев контрастируются временные узоры звуковых стимулов по интенсивности, частоте повторения посылок и серий, направленности. По-видимому, именно здесь происходят различение и опознание их видовой специфичности. Одновременно идёт фильтрация посторонних шумов и помех. В конце концов ассоциативные нейроны грудных ганглиев передают сообщения в высшие нервные центры слухового анализатора, расположенного в надглоточном ганглии.
Следовательно, обработка воспринятой акустической информации проходит по меньшей мере в три этапа: на уровне сенсорных нейронов рецептора, на уровне ассоциативных нейронов грудных ганглиев и, наконец, в головном мозге, где продолжаются опознание и локализация источника звука и формируется целостный образ раздражителя.
В разноголосом хоре насекомых, птиц и амфибий, среди шелеста листьев и трав самки кузнечиков рода Isophya не слышат песен сверчков, но отвечают на каждую звуковую посылку самцов своего вида одним-двумя характерными щелчками. Нередко к солирующему самцу присоединяются другие, подчиняя его песням ритмический рисунок своих трелей.
Терморецепция и гигрорецепция
Влияя на метаболические процессы, осмотические свойства цитоплазмы, свойства мембран и растворов, изменения температуры и влажности могли бы регистрироваться организмом непосредственно как изменения всех этих процессов и свойств. Более того, проведение импульсов возбуждения по нервам и синапсам и спонтанная активность нейронов также подвержены влияниям температуры и влажности, но не как рецепторных стимулов, а как экологических факторов, сравнимых по значению и характеру действия с влиянием корма, полового партнёра или хищника.
Вряд ли есть сомнения в том, что моментальная реакция на изменения температуры и влажности по сравнению с их регистрацией через изменения в метаболических процессах биологически целесообразна. Иными словами, вошедшему в огонь или в воду способность лишь к опосредованному восприятию такого рода факторов могла бы обойтись слишком дорого.
Вместе с тем не исключены возможные влияния влажности и температуры на рецепторные мембраны сенсорных нейронов механо- или хеморецепторов. Во всяком случае, насекомые имеют специализированные термо- и гигрорецепторы.
Трихоидные сенсиллы на антеннах таракана Periplaneta americana L. реагируют пятикратным увеличением частоты импульсов на понижение температуры на 1 °С. Дрессированные пчелы различают температуру посещаемых субстратов с точностью до 2 °С. Их фазотонические рецепторы имеют вид коротких волосков, погруженных в кугикулу (целоконические сенсиллы), и возбуждаются при понижении температуры. Однако сходные сенсиллы на антеннах хлебного жука-кузьки Anisoplia austriaca L. генерируют фазовые залпы импульсов лишь с повышением температуры, как, впрочем, и терморецепторы клопа Oncopeltus fasciatus L. На этом основании их именуют тепловыми в отличие от холодовых рецепторов пчелы и таракана.
Особой чувствительностью отличаются целоконические сенсиллы на кончиках антенн комара Aedes aegypti L. Располагая нейронами обоих типов, они генерируют фазовые ответы при изменении температуры на 0,05°С; при этом возбуждение теплового нейрона тормозит активность холодового и наоборот. Однако они не реагируют на источник инфракрасного излучения, воспринимая лишь конвекционное тепло — энергию молекул нагретого воздуха.
Удаление антенн у многих насекомых и регистрация потенциалов действия (ПД) нейронов некоторых чешуекрылых позволяют признать рецепторами влажности тонкостенные базиконические сенсиллы. Последние отличаются от трихоидных сенсилл тем, что имеют вид коротких конусов с закруглёнными вершинами. В ответ на струю влажного воздуха они генерируют импульсы, которые, однако, тормозятся сухим воздухом. Сходные реакции отмечены в некоторых нейронах механо-рецепторов кровососущих клопов родов Triatorna и Rhodnius, но дифференцировать их от возможных влияний влажности на механические (физические) свойства вспомогательных структур не представляется возможным.
Хеморецепция
Восприятие химических стимулов предполагает условное противопоставление дистантной и контактной хеморецепции. В первом случае стимуляторами служат летучие соединения, во втором — растворы этих соединений.
В обоих случаях источником стимула служит потенциальная энергия взаимного притяжения и отталкивания элементарных частиц на рецепторных мембранах. Вместе с тем очевидные различия в абсорбции газов и жидкостей определяют своеобразие вспомогательных структур, обслуживающих толстостенные вкусовые и тонкостенные обонятельные сенсиллы (рис. 123). Нередко и те и другие имеют облик трихоидных, базиконических и стилоконических сенсилл, но первые, как правило, несут одну вершинную пору, вторые — множество пор по всей поверхности рецептора.

Рисунок 123. Толстостенная (А) и тонкостенная (Б) сенсилла насекомых (по Gillot, 1980):
1 — сенсорные нейроны; 2 — кутикула; 3 — поры
Контактные хеморецепторы и вкусовой анализатор
Толстостенные вкусовые сенсиллы чаще всего сосредоточены на ротовых придатках и на лапках насекомых, реже — на их антеннах и яйцекладе. Обычно им сопутствуют 3–5 (реже — до 50) нейронов с дендритами, достигающими вершинной поры, которая способна замыкаться при механических раздражениях.
Несмотря на сравнительно небольшое число сенсорных элементов, вкусовые реакции и предпочтения насекомых далеко не однозначны. Например, в трёхнейронных лабеллярных сенсиллах мухи Phormia regina Mg. каждый нейрон выполняет определённые и специфические функции: один из них служит механорецептором, а два других дифференцированы на солевой (L-нейрон) и сахарный (S-нейрон). В более сложных четырёхнейронных сенсиллах наряду с ними присутствует W-нейрон, раздражаемый дистиллированной водой.
Стимуляция S-нейрона провоцирует положительную пищевую реакцию, основанную на лабеллярном рефлексе — развёртывании хоботка при контактах вкусовых рецепторов лапки с привлекающим пищевым субстратом. При раздражении L-нейрона пищевая реакция тормозится и насекомое стремится покинуть субстрат. При этом частота генерируемых импульсов пропорциональна силе воздействия, то есть концентрации растворов Сахаров или солей.
Отличаясь весьма низкой спонтанной активностью (1–4 импульса в секунду), оба нейрона обладают тормозящими воздействиями: раздражения солевого рецептора тормозят активность сахарного и наоборот.
Таким образом, нейроны контактных хеморецепторов насекомых позволяют различать привлекающие и отвергаемые вещества и более дробно дифференцировать их по концентрациям и свойствам.
Информация, поступающая в нервные центры анализатора, подвергается дальнейшей обработке, и возникающие на её основе вкусовые реакции весьма разнообразны. Насекомые дифференцируют четыре первичных вкусовых качества, различая сладкое, солёное, кислое и горькое, не уступая человеку по чувствительности к ним. Например, человек различает сахар в концентрации 2?10-2 моль/л, а пчёлы и бабочка-адмирал (Pyrameis atalanta L.) — в концентрации 6?10-2 и 10-1—10-2 моль/л соответственно.
Однако при длительном воздержании от сахаров бабочка-адмирал способна снизить порог чувствительности до 8?10-5 моль/л. Насекомые отличаются своеобразием вкусов. Например, безвкусная для пчёл лактоза оказалась умеренно сладкой для гусениц бабочек, воспринимающих с пищей существенно меньшее количество сахаров.
В отношении солей насекомым свойственны положительные пищевые реакции лишь на малые концентрации, но наиболее «солёны» для них катионы K, наименее — катионы Li.
Пороги чувствительности пчёл к кислым веществам — 10-3 моль/л. В отношении непереносимо горьких растворов хинина, ацетилсахарозы и настоев полыни насекомые малочувствительны.
Насекомые воспринимают вкус дистиллированной воды, которая для человека безвкусна; нередко они обнаруживают странное пристрастие к ядам (см. с. 78). Например, питающийся на зверобое листоед Chrysolina sp. имеет специальный рецептор, возбуждаемый только характерным для этого растения алкалоидом — гиперизидом.
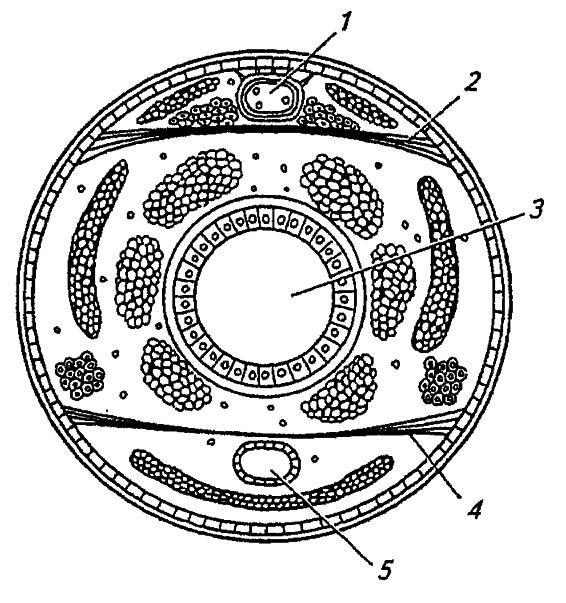
Рисунок 78. Общий план внутреннего строения насекомых (по Weber, 1966):
1 — спинной сосуд; 2 — дорзальная диафрагма; 3 — кишечник; 4 — вентральная диафрагма; 5 — нервная система
Дистантные хеморецепторы и обонятельный анализатор
Дистантные хеморецепторы, как правило, сосредоточены на антеннах; обычно их больше, чем контактных (до 60 000 у самцов бабочки Antheraea polyphemus L.), но все же насекомые значительно уступают по этому показателю млекопитающим. Например, у кролика имеется до 108 ольфакторных нейронов.
Среди дистантных хеморецепторов насекомых нередки трихоидные, базиконические, целоконические, а также плакоидные сенсиллы. У одних видов они рассеяны по всей поверхности антенн, у других — объединены группами под покровами в сложные органы обоняния.
Обонятельные сенсиллы обычно отличаются многочисленными порами, заполненными 10–20 трубочками, с которыми связаны ветвящиеся отростки дендритов. Назначение этой поровотрубчатой системы, видимо, состоит в активной абсорбции и транспортировке молекул ольфакторных раздражителей к рецепторным мембранам.
Пока остаётся неясным, каким образом немногие молекулы летучих соединений (феромонов), попадая на антенну, достигают рецепторов и вступают с ними во взаимодействие. Неясен и характер этого взаимодействия. По весьма популярной в своё время стереохимической теории обоняния обладающие определённой пространственной конфигурацией молекулы оседают на рецепторную мембрану, в которой имеются соответствующие этой конфигурации участки — лунки. На смену этой оригинальной, но плохо согласующейся с фактами гипотезе была выдвинута вибрационная теория, связывающая способность различать запахи с восприятием колебаний молекул, которые регистрируются по спектрам поглощения в инфракрасной области. Однако при замещении водорода тритием в молекулах особо привлекательных веществ они по-прежнему охотно воспринимались насекомыми, несмотря на резкое изменение спектров поглощения.
Тонкостенные обонятельные сенсиллы обычно содержат по нескольку нейронов; изредка их число достигает 40–60; при этом они специализируются на восприятии разных химических соединений.
В ответ на обонятельные стимулы нейроны генерируют импульсы, частоты которых зависят не только от концентрации веществ, но и от их привлекательности для насекомых.
Обычно при повышении концентрации привлекающих соединений частоты импульсов увеличиваются, а возрастание концентраций отвергаемых веществ приводит к её снижению.
Таким образом, на уровне дистантных хеморецепторов происходит первичная дифференциация раздражителей, чему в немалой степени содействуют два типа сенсорных нейронов: генералисты и специалисты. Первые из них отвечают на широкий набор разных летучих соединений, но малочувствительны, вторые реагируют на одно или несколько близких соединений и отличаются повышенной чувствительностью. Например, у жука-мертвоеда Necwphorus sp. нейроны этого типа обеспечивают регистрацию трупного запаха с расстояния до 90 м, у самцов шелкопряда Bombyx mori L. они позволяют различать половой аттрактант самок — бомбикол в концентрации 100 молекул на 1 см3. При этом для раздражения одного нейрона-специалиста достаточно одной молекулы, а для генерации возбуждения — не менее 1 молекулы в секунду в расчёте на поверхность обеих антенн. По образному замечанию Ж. А. Фабра, возможность рецепции самцом грушевой сатурнии запаха самки с расстояния 5–10 км (рекордный случай) можно сравнить с попыткой окрасить воды озера одной каплей красителя кармина.
Столь фантастичная чувствительность насекомых к специфическим запахам предполагает очень строгую дифференциацию ольфакторных стимулов. Аксоны соответствующих сенсорных нейронов переводят нервные импульсы в обонятельные доли дейтоцеребрума непосредственно, без синаптических переключений. По-видимому, существование изолированных путей для проведения возбуждения от нейрона-специалиста к ассоциативным центрам соответствует особому биологическому назначению ольфакторных стимулов.
Между тем по порогу чувствительности к некоторым запахам насекомых можно сравнить с человеком. Так, муха Phormia regina Mg. пары аммиака чувствует в концентрации 2?10-6 моль/л и пчела воспринимает отвратительно пахнущий скатол в концентрации 2,3?10-8 моль/л. Из предлагаемого пчеле набора 1 816 парных сочетаний запахов она при соответствующей дрессировке с успехом различает почти все из них (95,5 %).
Таким образом, насекомые различают разнообразнейшие запахи, среди которых одни принадлежат излюбленному корму, другие — субстратам для откладки яиц, третьи — ориентирам гнёзд или семьи. Среди них можно выделить запахи следов фуражирующих термитов или муравьёв, тревоги, потенциального полового партнёра и издаваемые пчелиной маткой для торможения развития яичников у рабочих особей.
Фоторецепция
В основе светоразличения лежит регистрация сенсорным нейроном фотоизомеризации зрительного пигмента — родопсина. Под воздействием поглощённого кванта света он преобразуется в изомер — метародопсин, который, восстанавливаясь в родопсин, обеспечивает многократное повторение элементарных зрительных актов.
Для восприятия зрительных стимулов насекомые располагают совершенными фоторецепторами — омматидиями, сгруппированными в сложные фасеточные глаза, дорзальными глазками и свойственными личинкам стеммами. В омматидии наряду с 4–12 униполярными нейронами, называемыми здесь ретинулярными клетками, выделяются вспомогательные структуры рецептора — светопреломляющий аппарат и аппарат пигментной изоляции. Первый из них образован шестигранной кугикулярной линзой роговицы, или хрусталиком (фасеткой), и лежащими под ней четырьмя земперовыми клетками, формирующими кристаллический конус. Снаружи каждая фасетка роговицы покрыта сетью мельчайших бугорков. Снижая отражение света и тем самым повышая прозрачность хрусталика, эта тончайшая сеть, видимая лишь под электронным микроскопом, придаёт ему свойства просветлённой оптики и устраняет блеск поверхности глаза. Клетки кристаллического конуса проводят в глубь омматидия тонкие нити световодов, что придаёт светопреломляющему аппарату глаз насекомых достоинства современных оптических приборов.
Сфокусированные собирательными линзами и проходящие по световодам лучи света достигают расположенных в глубине ретинулярных клеток. Группируясь вокруг оптической оси, как дольки апельсина, эти светочувствительные нейроны образуют направленные к центру микровилли — тончайшие микротрубочки, слагающиеся в рабдомер. Именно здесь совершаются элементарные акты свето-различения. В микровиллах каждого рабдомера, то есть оптически активной части светочувствительного нейрона, содержится до 107 молекул зрительного пигмента. Рабдомеры каждой ретинулярной клетки либо объединяются друг с другом в замкнутый рабдом, либо, например у мух, сохраняя обособленность, выстраиваются по стенкам центральной полости открытого рабдома (рис. 124).

Рисунок 124. Замкнутый (А) и открытый (Б) рабдом у насекомых (по Мазохину-Поршнякову, 1977)
В состав аппарата пигментной изоляции омматидия входят две корнеагенные (формирующие хрусталик) клетки, не менее четырёх ирисовых клеток и часто ещё несколько ретинальных пигментных клеток. В их цитоплазме рассеяны тёмные красноватые гранулы оммохромов, поглощающие почти все лучи видимого света, кроме красных, и более светлые гранулы птеринов, поглощающие ультрафиолетовые лучи. Образуя светонепроницаемый чехол вокруг светочувствительных элементов глаза у дневных насекомых, эти клетки у сумеречных и ночных форм способны концентрировать гранулы пигмента близ хрусталика, открывая рабдом ретинулярных клеток для засветки боковыми лучами соседних омматидиев. Сокращая остроту восприятия, это явление (ретиномоторная реакция) многократно увеличивает светосилу глаза. При низкой освещённости, в сумерках, такое повышение светосилы очень важно. Однако качество изображения, создаваемого оптическими структурами, не имеет большого значения, так как обозреваемый насекомыми объект воспринимается каждым омматидием по отдельности, как мозаика пятен разной яркости.
В соответствии с отмеченными различиями свойств и набора пигментных клеток можно противопоставить аппозиционным глазам дневных форм суперпозиционные сумеречных и ночных насекомых.
Различаясь, кроме того, длиной рабдомов, эти два типа глаз предполагают либо сложение (аппозицию) воспринимаемого изображения из отдельных улавливаемых омматидием пятен, либо наложение (суперпозицию) этих пятен друг на друга. Способность пигментных клеток суперпозиционного омматидия рассредоточиваться по всей длине рабдома приводит его в функционально аппозиционное состояние (рис. 125), что происходит всякий раз при избытке освещения днем. Рассредоточение пигмента завершается в считанные минуты, тогда как для приспособления глаза к ночному видению требуется от 10 до 50 мин.
У двукрылых, отличающихся открытым рабдомом, ориентация отдельных рабдомеров омматидия такова, что все они «смотрят» в разные точки пространства, однако те же самые точки воспринимаются шестью рабдомерами, принадлежащими разным омматидиям. Иннервирующие их волокна сходятся в одном нервном центре, поэтому сигналы ретинулярных клеток разных омматидиев суммируются в оптических долях протоцеребрума. Таким образом, сохраняя преимущества аппозиционного видения, нейросуперпозиционные глаза мух обладают повышенной светосилой. Однако по сравнению с суперпозиционными глазами ночных бабочек, светосила которых превосходит аналогичный показатель аппозиционных глаз в десятки и сотни тысяч раз, нейросуперпозиционные глаза гораздо менее чувствительны в сумерках или ночью.
Глаза дневных форм насекомых также способны приспосабливаться к сумеречному освещению, но только за счёт нарушения равновесия фотоизомеризации зрительного пигмента и изменения коэффициентов преломления ретинулярных клеток.
При поглощении рабдомерами фотона света они генерируют миниатюрные импульсы, видимо, связанные с выделением медиаторов в ретинулярные клетки, которые в ответ на это генерируют постоянно нарастающий потенциал деполяризации мембраны. Такого рода безимпульсные сигналы характерны для светочувствительных элементов, что отличает их от сенсорных нейронов других типов. Достигая оптических долей протоцеребрума, эти сигналы переключаются на ассоциативные нейроны, связывающие друг с другом разные ганглиозные массы. Здесь (см. рис. 116) выделяются четыре отдела и нервные волокна большинства омматидиев скрещиваются в оптической пластинке.

Рисунок 116. Головной мозг насекомых (по Romoser, 1981):
1 — оптическая пластинка; 2 — медуллярные пластинки; 3 — оцеллярные нервы; 4 — протоцеребральный мост; 5, 6 —соответственно грибовидное и центральное тело; 7 — дейтоцеребрум; 8 — тритоцеребрум
При замкнутом рабдоме волокна объединяют аксоны ретинулярных клеток одного омматидия, при открытом рабдоме — аксоны разных омматидиев, рабдомеры которых имеют совпадающую ориентацию. Вместе с тем отдельные сенсорные нейроны передают воспринимаемую информацию сразу в медуллярную пластинку, связанную с оптической пластинкой нервными волокнами зрительного перекреста. Дальнейшие пути проведения нервного возбуждения ещё не исследованы, но оптические волокна отмечены и в центральных областях протоцеребрума, и в стебельчатых телах.
Важнейшие рабочие элементы зрительных центров — специализированные нейроны, которые представляют собой детекторы, избирательно реагирующие на параметры зрительных стимулов: их размеры, форму, окраску и характер движения. Нередко выделяются только те из них, которые движутся в строго определённом, предпочитаемом для восприятия направлении. Движущиеся иначе тормозят импульсную активность и, по-видимому, не воспринимаются насекомыми.
Наиболее простые детекторы — нейроны тревоги, реагирующие на затенение части глаза и любые перемещения объектов в поле зрения. Генерируемые ими импульсы обусловливают включение рефлекторной реакции бегства. Несколько сложнее организованы детекторы признаков полового партнёра или направленности движений, в частности детектор движущегося пятна у стрекоз, включающий реакцию преследования.
Своеобразен рефлекторный механизм латерального торможения, проявляющийся в том, что активность омматидиев зависит не только от силы стимула, но и от активности соседних омматидиев. Усиливая перепады в освещённости созерцаемых объектов, эти механизмы подавляют оптически вредные последствия перекрывания полей зрения соседними омматидиями, не препятствуя, однако, стереоскопичности видения. Отметим, что сходные принципы реализованы в декодирующих устройствах реставраторов и криминалистов, используемых для расшифровки нечётких фотографий.
Распределение омматидиев по сфере сложного глаза и их численность, достигающая 12 000–17 000 у бабочек и 10 000–28 000 у стрекоз, придаёт зрению насекомых некоторые особые свойства.
Очевидно, что чем ближе сведены оптические оси омматидиев, чем меньше между ними угол, тем выше острота зрения — способность к различению двух сближенных между собой точек. У насекомых она в 3 раза выше, чем у человека (30 и 100 мкм соответственно), но вместе с тем при удалении объекта от глаза острота зрения резко сокращается — насекомые близоруки.
Кроме того, при малейших смещениях объекта его восприятие сразу же переходит к другим, соседним омматидиям, сообщая насекомым особую чувствительность к движущимся стимулам. Наряду с весьма малой инерционностью рецепторов это придаёт глазу способность к различению быстро мелькающих предметов. Если для человека слитное изображение на экране возникает при смене 16 кадров в секунду, то для насекомого — 250–300. Вполне понятно, что это свойство необходимо при скоростном полете среди растений. Ни человек, ни насекомое вообще не видят объектов, неподвижных в отношении сетчатки глаза. Постоянный тремор глазодвигательных мышц человека — одно из непременных условий видения — противопоставляется неподвижности глаз насекомых, которые для выполнения этого условия вынуждены вибрировать всем телом. Обычно при громадном числе фасеток это не требует больших затрат энергии, но если их число ограничено, как, например, у гусениц бабочек, то необходимы более интенсивные движения тела. Быть может, именно этим объясняется особенность гусениц отрываться время от времени от приёма пищи, приподнимая и раскачивая переднюю часть тела.
Сходным образом объясняется различение насекомыми формы объектов и предпочтение ими тех, чей контур более рассечён. Кроме того, уже доказана стереоскопичность зрения насекомых, обеспечивающая восприятие объёмов тел и глубины пространства, а также способность к восприятию поляризации света. Не только голубое небо, но и поверхность водной глади, и покрытых восковым налётом листьев растений, и покровов насекомых, поляризуя свет, придают им неразличаемые человеком блики и тени.
В ещё большей мере обогащает зрительные восприятия насекомых способность к цветовому зрению, доказанная для многих из них. Они не только могут различать спектральный состав света, но и воспринимают, как и человек, естественную окраску предметов независимо от цвета освещения. Среди исследованных в этом отношении насекомых цветослепотой страдали лишь пещерные формы, термиты и большой мучной хрущак (Tenebrio molitor L.).
Способность к цветоразличению определяется по меньшей мере двумя светоприёмниками с различной спектральной чувствительностью. У человека их три, у пчёл и многих дневных бабочек — также три, у других насекомых — обычно только два. Один из них, как правило, настроен на восприятие ультрафиолетовых лучей и обладает повышенной чувствительностью, отличая, например, жёлтый цвет от жёлтого с 1 % примеси ультрафиолетового света. На рисунке 126 приведена схема различной спектральной чувствительности глаз человека, пчелы и мухи.
Ощущения, испытываемые человеком при восприятии различных стимулов, всегда субъективны. Поэтому, если пчелы, например, различают жёлтый и зелёный цвета, мы не знаем, как выглядят эти цвета для них, как они ощущаются ими. Кроме того, многие насекомые обладают врождённой предрасположенностью, например, к синим и пурпурным (бабочка-парусник Papillo troilus L.) или жёлтым цветам (мухи-журчалки рода Eristalis, многие тли) и не отличают белый цвет от смеси жёлтого с сине-фиолетовым.
Однако особое своеобразие цветовым восприятиям придает ультрафиолетовая окраска. Одноцветный для человека самец бабочки-лимонницы Gonepteryx rhamni L. воспринимается его самкой как двуцветный, так как его передние крылья сильно отражают ультрафиолетовые лучи. Однотонно-белые капустная белянка (Pieris brassicae L.) или бабочка-боярышница (Aporia crataegi L.) для насекомых весьма различны по цвету и имеют явные признаки половой принадлежности.
Многие жёлтые или белые цветки различаются насекомыми по ориентирам нектарников, а красный мак для невосприимчивых к красным лучам мух «сияет» лишь одним ультрафиолетовым «цветом».
Наряду с омматидиями сложных глаз в восприятии зрительных стимулов принимают участие дорзальные глазки, а у лишённых сложных глаз личинок они замещены стеммами. И те и другие значительно проще по устройству, но отличаются некоторыми своеобразными функциями.
Светопреломляющая кутикулярная линза дорзальных глазков отграничена слоем мелких корнеагенных клеток (рис. 127) от глазного бокала, в котором сосредоточены светочувствительные ретинальные клетки часто вперемежку с пигментными. Отходящие от глазков аксоны сенсорных клеток достигают расположенных рядом оцеллярных ганглиев, переводя воспринятую зрительную информацию к ассоциативным нейронам.

Рисунок 127. Строение дорзального глазка насекомых (по Шванвичу, 1949):
1 — нервы; 2 — пигментные клетки; 3 — хрусталик; 4, 5 — соответственно корнеагенные и ретикулярные клетки
Таким образом, ганглии глазков находятся вне протоцеребрума, но тесно связаны с ним толстыми оцеллярными нервами (см. рис. 116). Глазки регистрируют изменения освещённости и наступление темноты, различают цвета и поляризацию света, воспринимают движение крупных объектов и при этом обладают повышенной светосилой. Кроме того, поступающие от них сигналы влияют на активность головного мозга, который получает высокое тоническое возбуждение даже в темноте, регулируют направленность движения к источнику света и ритмику биений спинного сосуда. Однако некоторые весьма активные насекомые лишены развитых глазков.
Рисунок 116. Головной мозг насекомых (по Romoser, 1981):
1 — оптическая пластинка; 2 — медуллярные пластинки; 3 — оцеллярные нервы; 4 — протоцеребральный мост; 5, 6 —соответственно грибовидное и центральное тело; 7 — дейтоцеребрум; 8 — тритоцеребрум
Личиночные стеммы разнообразны по строению. Более примитивные из них напоминают дорзальные глазки, более совершенные — отдельные омматидии. Отличаясь высокой светосилой, они способны к цветоразличению и восприятию поляризации света.
Обобщение зрительных образов
О способности насекомых к дрессировке, запоминанию и обобщению зрительных образов можно судить по результатам экспериментов, в которых использованы условно-рефлекторные реакции на определённые стимулы.
Предлагая пчёлам корм, расположенный на окрашенном столике, нетрудно уже через 2–3 прилёта приучить их к определённому цвету как к символу пищи. Формированию условного рефлекса на двуцветные стимулы соответствует предпочтение и иных двуцветных сочетаний, даже если каждый из избранных цветов ранее не использовали при дрессировке.
Наиболее поразительные демонстрации способностей насекомых принадлежат К. Фришу, открывшему танец пчелы-разведчицы. Вернувшись в улей, она сообщает членам своей семьи о найденном ею корме и о том, где фуражирам его следует искать. Пробегая некоторое расстояние по стенке улья, пчела-разведчица виляет брюшком и, сворачивая направо и налево, описывает круги (рис. 128). Ориентация траектории прямого пробега в отношении направления силы тяжести точно соответствует направлению полёта при ориентации по Солнцу, а частота виляний брюшка — расстоянию до источника корма. Если танец происходит на горизонтальной плоскости камеры с верхним смотровым окошком, то ориентация прямого пробега соответствует направлению поиска, но только тогда, когда небо не затянуто облаками. В противном случае танец становится ненаправленным. Стоит пчеле увидеть хотя бы кусочек голубого неба в разрыве облаков, танец вновь становится ориентированным.

Рисунок 128. Танец пчелы-разведчицы (по Frish, 1962)
Таким образом, наряду с расшифровкой смысла танца пчелы К. Фриш открыл способность насекомых к восприятию поляризации света и использование этой способности для наведения на цель. В сущности это те же способы ориентации, которыми пользуется человек при движении по азимуту. Очевидно, что компасом насекомых может быть Солнце или направление поляризации лучей света голубого неба.
Преобразование ориентиров, в данном случае угла направления на Солнце в вертикальный угол, образованный траекторией прямого пробега в отношении силы гравитации, свидетельствует о более глубоких способностях насекомого. Однако даже эти способности все же недостаточны для ориентации многочасовых ночных перелётов и миграций на тысячи километров. Возникающая необходимость коррекции направления полёта по астрономическим ориентирам требует не только ощущения времени, но и синхронизации внутренних биологических часов организма с суточным движением Солнца на разных широтах и в различных часовых поясах. Демонстрируя качества опытных навигаторов, насекомые справляются и с этими, далеко не простыми задачами. Возможно, они регистрируют геомагнитное поле, но для однозначного ответа на эти вопросы требуются более глубокие исследования.
Поведение
Воспринятые насекомыми стимулы служат основой для формирования целесообразных актов поведения как результата сочетания врождённых рефлексов, инстинктов и приобретённого опыта.
Врождённые, или безусловные, рефлексы проявляются как однозначные ответы на раздражители. Примеры их — описанные ранее тарзальный и лабеллярный рефлексы, рефлекс движущегося пятна, включающий реакцию преследования у стрекоз и бросок богомола. Серии безусловных рефлексов определяют стереотипы пищевого и сексуального поведения насекомых, поведения при выборе мест откладки яиц и инстинкты при строительстве гнёзд и домиков, при завивке коконов и тому подобного.
Инстинкты и стереотипы поведения представляют собой цепи последовательных рефлексов, которые возникают в ответ на изменения физиологического состояния организма. В этом случае их побудительными мотивами могут быть и голодание, и созревание половых продуктов или изменения в концентрации гормонов и других физиологически активных веществ, соответствующие достижению определённых фаз и стадий развития.
Сходную нейрофизиологическую природу имеют таксисы и тропизмы. Их следует противопоставить друг другу как алгоритмы наведения на цель и способы ориентации вне зависимости от природы ориентирующего стимула (таксисы) и как собственные отношения к этим стимулам, связанные с внутренней мотивацией их выбора (тропизмы). В соответствии с набором стимулов и отношением к ним насекомых различают положительные и отрицательные фото-, гео-, хемо-, гигро-, термотропизмы и так далее, подразумевая положительные и отрицательные реакции на источники света, гравитацию, источники химических стимулов, влажности и тепла.
Механизмы формирования тропизмов до сих пор остаются неясными, но их биологический смысл часто вполне очевиден. Известны многочисленные примеры использования тропизмов насекомых (как и рефлексов, и инстинктов) в практике защиты от них растений, в частности при наложении ловчих поясов на штамбы плодовых деревьев (отрицательный геотропизм гусениц плодожорок), при уничтожении растительных остатков, в которых скрываются от света многие вредители, при организации вылова насекомых различными светоловушками и приманками с феромонами.
Что касается таксисов, то наиболее явные из них — это фобо-, клино-, тропо-, тело- и менотаксис.
Фоботаксис, иногда называемый методом проб и ошибок, представляет собой относительно примитивный, но и наиболее общий алгоритм поведения в экстремальных условиях, нередко угрожающих самому существованию организма. Он проявляется в замедлении или ускорении движений под влиянием стимула и в частоте изменений направления движения. Например, при повышенной температуре многие насекомые начинают быстро бегать, постоянно меняя направление движения, при этом вероятность выхода за пределы зоны опасности возрастает. На основе фоботаксиса можно объяснить формирование скоплений насекомых в местах с благоприятными для них режимами влажности, освещённости, температуры.
Клинотаксис определяет целенаправленность движения насекомого при последовательном выборе уровней возбуждения соответствующих рецепторов. Например, личинки комнатной мухи, отличающиеся отрицательным фототропизмом, раскачивают переднюю часть тела с расположенными на ней фоторецепторами, избирая направление движения, при котором рецепторы возбуждаются минимально.
Тропотаксис представляет собой более совершенный алгоритм наведения на источник стимула, основанный на достижении равновесия в возбуждении симметричных рецепторов. Например, медоносная пчела, расставив в стороны антенны, легко воспринимает пахучую приманку, расположенную в одном из колен Y-образной камеры. Если её антенны закрепить на лбу в перекрещённом состоянии, то она всегда направляется в противоположное колено камеры на основе тропотаксиса. При нарушении симметрии рецепторов, закрашивая, например, один глаз, мы вынуждаем насекомое к постоянным манежным (круговым) движениям даже в параллельном пучке света.
Телотаксис предполагает наведение на стимул за счёт того, что его воспринимают рецепторы, направленные вперёд. При этом даже ослеплённое на один глаз насекомое сохраняет верную ориентацию, не совершая манежных движений. По-видимому, целенаправленный бросок богомола основывается на телотаксисе, проявления которого бывает трудно отличить от проявлений тропотаксиса и менотаксиса.
Менотаксис своеобразен в том отношении, что наведение посредством его на цель предполагает фиксацию некоторого угла между направлением движения и ориентирующим стимулом. При этом избранные ориентиры могут быть несущественными для насекомого, не всегда строго фиксированными по положению в отношении истинной цели. В частности, привлечённые на свет бабочки демонстрируют менотаксис, который сравним со способами ориентации по азимуту.
Таким образом, многие аспекты поведения насекомых расшифровываются на основе врождённых, безусловно-рефлекторных реакций. Однако вопреки мнению Ж. Фабра насекомые не являются рабами своих инстинктов, своего рода автоматами, действующими по заранее разработанной программе. Приведённые ранее примеры формирования условных рефлексов у насекомых, их способностей к обобщению и дрессировке придают поведению гибкость, необходимую в переменчивых условиях существования. Известны примеры, когда формирующиеся условные рефлексы тормозят проявления инстинктов и тропизмов. Подвергая тараканов воздействию слабых электрических разрядов всякий раз, когда из двух предлагаемых им камер — освещённой и затемнённой — они выбирают последнюю, можно вскоре подавить проявления отрицательного фототропизма и приучить их к существованию в непривычных условиях (при освещении).
Итак, сенсорные системы насекомых, различающиеся по сложности организации вспомогательных структур, служат проводниками информационных стимулов внешнего мира. Вместе с тем они регистрируют состояние внутренней среды организма.
Энергия воспринятых стимулов, преобразованная сенсорными нейронами в специфические процессы нервного возбуждения, подвергается дальнейшей обработке и фильтрации в центральных отделах анализаторов. Формируя целостный образ раздражителей на основе врождённых реакций и приобретённого опыта, насекомые обнаруживают явные способности к дрессировке, запоминанию и обобщению, не уступая в этом отношении позвоночным животным. При этом насекомые сохраняют своеобразие, несмотря на все сходства целесообразных реакций и актов поведения с таковыми позвоночных животных, противопоставляются им как существа, обитающие в мире иных масштабов и стимулов.

