Морфофункциональные основы размножения насекомых
Воспроизводство потомков, обеспечивающее продолжение рода, нередко противоречит метаболическим потребностям индивидов. Израсходовав резервы, необходимые для поддержания своей собственной жизнедеятельности, на продукцию спермиев и яиц, многие насекомые требуют дополнительного питания для восстановления этих резервов или гибнут вскоре после размножения.
Мужские половые органы и сперматогенез
Мужские половые органы насекомых образованы парными семенниками (рис. 130), в которых формируются сперматозоиды, придаточными железами, выделяющими секреты, и семяпроводами.
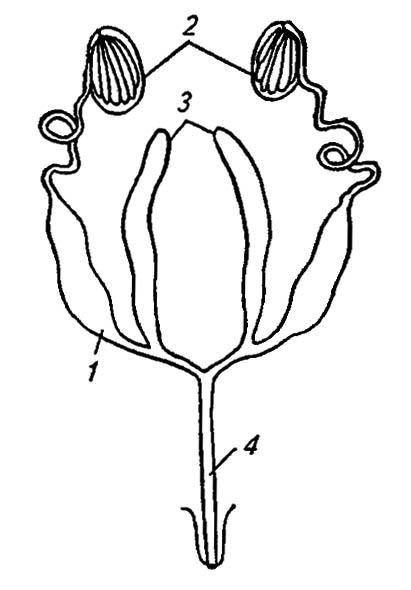
Рисунок 130. Строение половых органов самцов (по Romoser, 1981):
1 — семенной пузырёк; 2 — семенники; 3 — придаточные железы; 4 — семяизвергательный канал
Каждый семенник состоит из нескольких семенных фолликулов, расположенных посегментно у примитивных форм и объединённых в компактный орган с собственной оболочкой у более совершенных форм. В некоторых случаях происходит вторичное сокращение числа фолликулов (до одного у двукрылых, блох и сеноедов) или их увеличение (до 70–80 у прямокрылых и тараканов). Реже, например у веснянок и бабочек, правый и левый семенники срастаются друг с другом в непарное образование с двумя отходящими от него семяпроводами.
В ограниченной эпителием полости фолликула выделяются зоны последовательного развития половых клеток (рис. 131). В зоне размножения, или гермарии, сосредоточены интенсивно делящиеся диплоидные клетки первичных сперматогониев. Здесь же находится крупная апикальная клетка или группа клеток апикальной розетки, которые транспортируют к половым клеткам вещества, необходимые для их роста и развития.
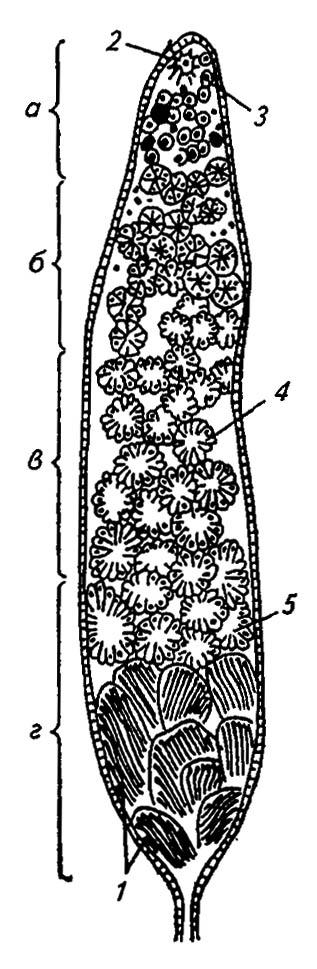
Рисунок 131. Фазы сперматогенеза в семеннике (по Gillot, 1980):
а — гермарий; б, в, г — соответственно зоны роста, созревания, трансформации; 1 — сперматозоиды; 2 — апикальная клетка; 3 — сперматогонии; 4 — сперматоциты; 5 — сперматиды
Потомки первичных сперматогониев — многочисленные вторичные сперматогонии, сохраняя плазматические связи друг с другом, оформляются в тонкостенные семенные цисты. Пройдя ряд синхронных митозов, они преобразуются в сперматоциты первого порядка, число которых в каждой цисте соответствует геометрической прогрессии: 32 у червецов, 64 у мух, 128 у ручейников и 256 у бабочек.
Проходя в зону роста и затем в зону созревания, каждый сперматоцит первого порядка подвергается редукционному делению и даёт начало четырём гаплоидным потомкам — сперматоцитам второго порядка. Последние, приобретая облик сперматид, теряют оболочку цисты и постепенно преобразуются в сперматозоиды в зоне трансформации. Все процессы сперматогенеза обычно завершаются у нимф и куколок до окрыления, и половозрелые самцы уже не пополняют расходуемые запасы половых клеток.
Зрелые сперматозоиды насекомых имеют веретеновидную головку, покрытую шапочкой акросомы, и удлинённый жгутик, образованный микрофибриллами осевого филамента (рис. 132). Сохраняя крупные митохондрии и центриоли, они теряют свойственные сперматидам рибосомы, структуры аппарата Гольджи и почти всю цитоплазму. Представляя собой ядро, снабжённое мощным, но кратковременно действующим двигательным аппаратом, сперматозоид специализирован в направлении поиска и осеменения яйца. Позднее, при слиянии с яйцеклеткой, он отбрасывает жгутик и в виде гаплоидного ядра мужского пронуклеуса участвует в образовании синкариона — диплоидного ядра зиготы.
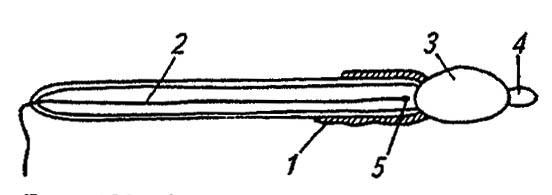
Рисунок 132. Строение сперматозоида (по Romoser, 1981):
1 — митохондрии; 2 — осевой филамент; 3 — головка; 4 — акросома; 5 — центриоль
Сформированные в фолликулах сперматозоиды проходят по семяпротокам в семяпроводы и остаются неподвижными в обширных полостях семенных пузырьков. Здесь они поддерживают жизнедеятельность за счёт секретируемых эпителием семяпроводов выделений и в момент копуляции пассивно выносятся волнами перистальтики в семяизвергательный канал.
Придаточные железы семенника имеют вид парных извитых трубочек, реже они отсутствуют (у двукрылых и первичнобескрылых) или объединены в крупный грибовидный орган (тараканы). Выделяемые ими секреты активируют движение сперматозоидов и вместе с тем побуждают самку к ускоренному формированию яиц. Нередко эти секреты содействуют развитию сексуальной восприимчивости, а также, заполняя половые пути самки, предотвращают повторные спаривания с другими самцами. Кроме того, выделения придаточных желез изредка поедаются самками или используются для образования сперматофоров — заполненных сперматозоидами флаконов (рис. 133).
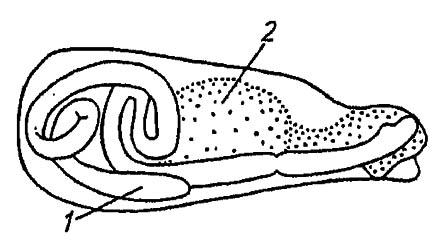
Рисунок 133. Строение сперматофора (по Romoser, 1981):
1 — семенной флакон; 2 — белковый секрет
Примитивные насекомые откладывают сперматофоры на субстрат, и самка подбирает их своими половыми придатками нередко при участии самца, подталкивающего её к ним. У многих сверчков и кузнечиков сперматофоры образуются до спаривания, у саранчовых — в момент спаривания, у тараканов и бабочек они формируются в половых путях самки из введённых самцом секретов.
Возможные пути регуляции деятельности придаточных желез представлены на рисунке 134. Добавим, что экдизон ограниченно стимулирует митотическую активность сперматогониев, а ювенильный гормон тормозит.

Рисунок 134. Возможные пути регуляции деятельности придаточных желез при формировании сперматофоров. Пунктирные линии — предполагаемые пути регуляции (по Gillot, 1980)
Женские половые органы и оогенез
Парные яичники насекомых образованы серией овариол — яйцевых трубочек, в которых проходят все важнейшие процессы формирования яиц (рис. 135). Закладывающиеся у эмбриона в виде посешентно расположенных парных зачатков овариолы сохраняют сегментарную принадлежность лишь у самых примитивных насекомых (рис. 136) и обычно утрачивают её при вторичной полимеризации зачатков. В исходном состоянии их число ограничено 5–7 парами в соответствии с набором половых сегментов (второй—девятый сегменты брюшка), в чем прослеживается сходство с кольчатыми червями, половые органы которых развиваются в сегментах задней половины тела.
Рисунок 134. Возможные пути регуляции деятельности придаточных желез при формировании сперматофоров. Пунктирные линии — предполагаемые пути регуляции (по Gillot, 1980)
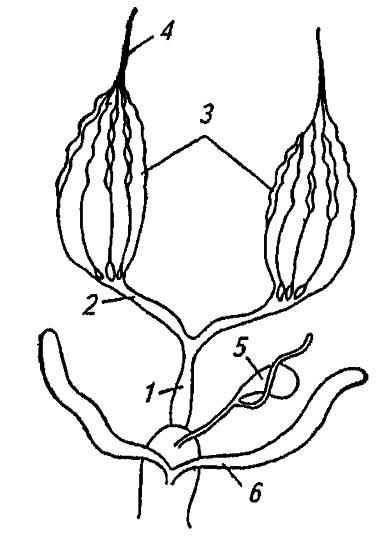
Рисунок 135. Строение половых органов самки (по Romoser, 1981):
1, 2 — соответственно медиальный и латеральный яйцевод; 3 — овариолы; 4 — терминальный филамент; 5 — семяприёмник; 6 — придаточные железы
Следовательно, весьма ограниченная плодовитость примитивных насекомых связывается с ограниченным числом сегментарных зачатков овариол, но если для кольчатых червей один из способов повышения плодовитости — рост их тела в длину, то для насекомых этот путь неприемлем.
Рассмотрим строение отдельной овариолы (рис. 137). Снаружи каждая яйцевая трубочка облечена слоем мышц, эпителием и соединительнотканной оболочкой (туникой). Её вершина продолжается в бесструктурный филамент, и все филаменты овариол правого и левого яичников, соединяясь друг с другом, закрепляются в дорзальной диафрагме. Обычно в пределах овариолы выделяют концевую камеру — гермарий и образованный серией фолликулов вителлярий. В гермарии сосредоточены первичные оогонии, митотическое размножение которых пополняет набор расходуемых гамет, а также редкие префолликулярные клетки. Начинающиеся здесь процессы оогенеза приводят к формированию вторичных оогониев, судьба которых в овариолах разных типов различна.
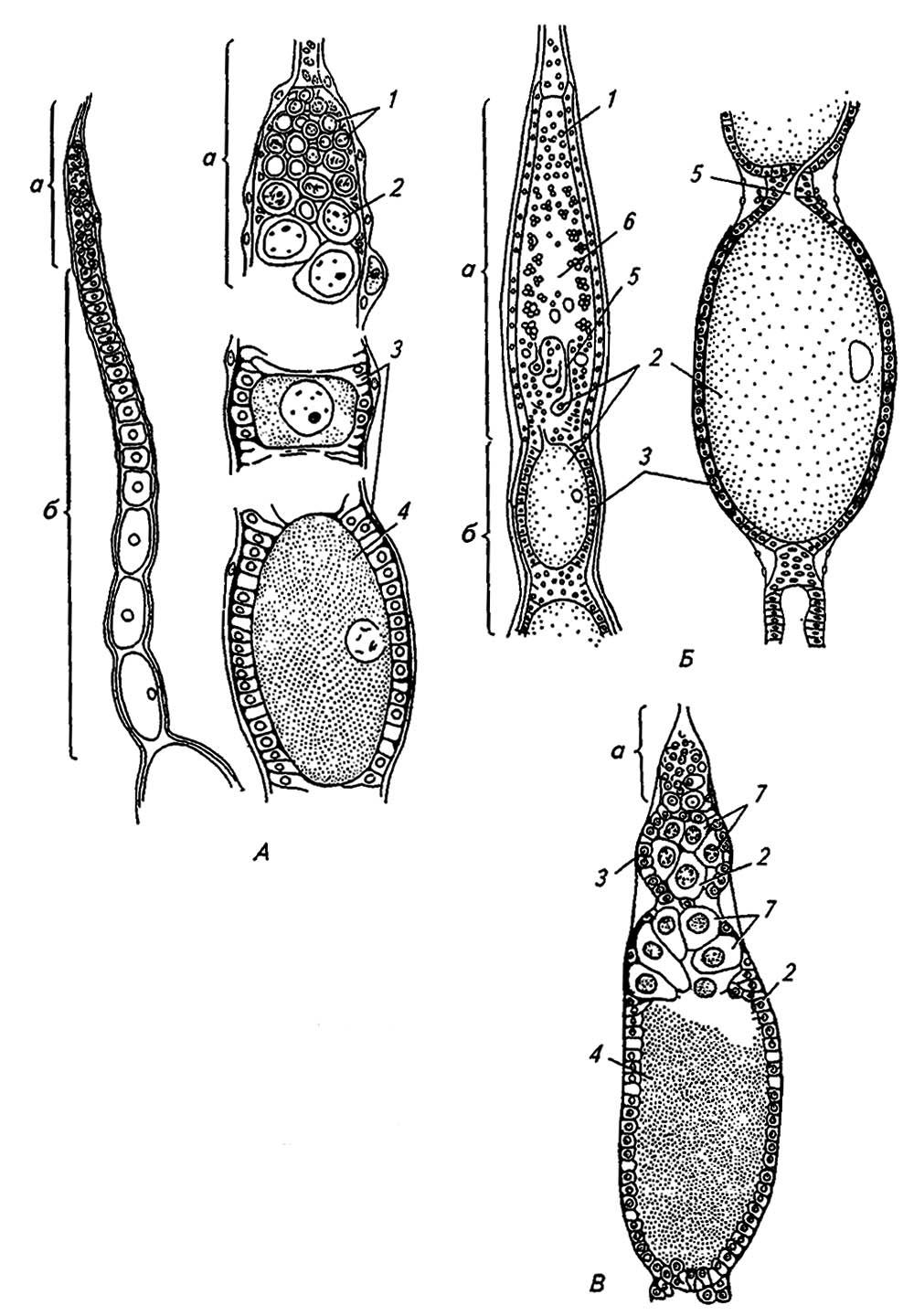
Рисунок 137. Строение паноистической (А), телотрофической (Б) и политрофической (В) овариолы у насекомых:
а — гермарий; б — вителлярий; 1 — оогонии; 2 — ооцит; 3 — фолликулярные клетки; 4 — желток; 5 — желточные тяжи; 6 — трофический синцитий; 7 — трофоциты
В примитивных паноистических овариолах, свойственных щетинохвосткам, подёнкам, стрекозам, всем ортоптероидным насекомым, исключая уховерток, а также трипсам, сеноедам, веснянкам, некоторым скорпионовым мухам, блохам и, вероятно, вислокрылкам, оогонии расходуются на образование яиц и в редких случаях на пополнение набора префолликулярных клеток. Преобразуясь в ооциты, они накапливают цитоплазму (фаза превителлогенеза) и, увеличиваясь в размерах, сосредоточиваются у основания гермария. Затем они проходят в вителлярий, где окружаются фолликулярными клетками и начинают накапливать желток (фаза вителлогенеза).
Фолликулярные клетки, образующиеся из префолликулярных клеток гермария, выстраиваются в эпителий и транспортируют из гемолимфы необходимые для синтеза желтка соединения. В это же время диплоидное ядро ооцита теряет оболочку и, принимая облик зародышевого пузырька, включается в синтез желточных гранул. Однако в отличие от ядер обычных секреторных клеток ядро ооцита никогда не претерпевает эндомитозов и полиплоидизации, интенсифицирующих процессы синтеза, в противном случае оно утратило бы роль хранителя генетической информации материнского организма, обрекая последующие поколения на вырождение и уродства. Пополнение набора матриц, необходимых для синтеза специфических протеинов, происходит за счёт своеобразной амплификации (умножения числа) генов в ядре ооцита. Однако и это средство не радикальное в решении проблемы интенсификации синтеза, так как процессы вителлогенеза в примитивных паноистических овариолах весьма продолжительны и постепенны. Столь же продолжительна и постепенна откладка сформированных яиц, число которых ограничено числом яйцевых трубочек яичника.
Более радикальное средство повышения плодовитости всех перечисленных выше насекомых — полимеризация зачатков овариол, число которых у подёнок достигает нескольких сотен, а у царицы термитов — нескольких тысяч. Другие насекомые избрали иные пути совершенствования оогенеза, которые предопределены ранней дифференциацией вторичных оогониев на ооциты и абортивные питающие клетки — трофоциты.
В мероистических овариолах размножение первичных оогониев завершается до окрыления и половозрелые самки лишь расходуют накопленные в гермарии запасы половых клеток. Вторичные оогонии, именуемые здесь цистобластами, после ряда синхронных делений формируют группы клеток — цистоцитов, связанных друг с другом цитоплазматическими тяжами — фусомами. При этом цистоциты разных поколений различаются числом фусом, которое зависит от числа делений, что предопределяет их дальнейшую судьбу. У некоторых жуков фактором дифференциации цистоцитов служит хроматическое тельце, проникающее лишь в одну из делящихся клеток — будущий ооцит. Все остальные клетки становятся позднее трофоцитами и либо сливаются друг с другом в единый трофический синцитий гермария, либо сопровождают ооцит в фолликулах витгеллярия. В телотрофических овариолах, свойственных уховёрткам, пухоедам, вшам, плотоядным жукам, сетчатокрылым, многим скорпионницам, перепончатокрылым, ручейникам, чешуекрылым, двукрылым и немногим блохам, общее число трофоцитов, сопутствующих единственному ооциту, варьирует от 1 (комары-звонцы) до 127 (жужелица Carabus violacens L.).
Пройдя фазы превителлогенеза в пределах гермария и в первых фолликулах вителлярия, ооцит приступает к вителлогенезу, а ядра трофоцитов, подвергаясь эндомитозам и полиплоидизации, принимают на себя основную работу по синтезу предшественников желтка. Транспортируя по фусомам не только относительно простые соединения, но и нуклеиновые кислоты, а также отдельные органеллы (митохондрии и рибосомы), трофоциты избавляют ооцит от многих функций, но все же окончательный синтез и распределение желточных запасов контролируются его ядром.
Роль фолликулярных клеток в мероистических овариолах более ограничена, чем в паноистических овариолах. Поставляя некоторое количество питательных веществ из гемолимфы, они вместе с тем сохраняют одну из основных своих функций — секрецию оболочки яйца — хориона и участвуют в резорбции погибших или по каким-либо причинам не развившихся ооцитов.
У некоторых паразитических и живородящих насекомых вителлярий редуцирован или отсутствует. Это объясняется следующим: у паразитических насекомых яйца не содержат желтка, отсутствие которого компенсируется специальными приспособлениями, позволяющими зародышу усваивать ткани и соки поражаемого организма; при живорождении материнский организм обеспечивает развивающегося потомка не через желток яиц, а непосредственно питательными веществами гемолимфы. При гемоцельном живорождении, заканчивающемся гибелью самки, развивающаяся личинка поглощает её ткани, что весьма напоминает паразитизм. Во всяком случае, и при живорождении, и при паразитизме отпадает необходимость в образовании желтка и, следовательно, в том отделе овариолы, где происходят процессы вителлогенеза.
Существование специализированных трофоцитов намного облегчает развитие яйцеклетки, но требует мобилизации всех метаболических резервов для интенсивной, хотя и скоротечной деятельности. Формируя яйца одновременно во всех овариолах яичника, самка способна откладывать последовательные партии яиц с интервалами, необходимыми для восстановления израсходованных запасов пластических и резервных веществ. Продуцирование яиц становится, таким образом, важнейшим событием в её жизни, разделённой на последовательные гонотрофические циклы (лат. gonos — размножение, trophos — питание). У кровососущих комаров закономерные чередования приёма пищи и откладки яиц достигают состояния совершенной гонотрофической гармонии: получив полную порцию крови, они способны к развитию полной партии яиц, и лишь у видов с автогенным развитием яичников первая партия яиц формируется до крово-сосания в ущерб потребностям самого насекомого. За лето самка малярийного комара Anopheles maculipennis Mg. испытывает до пяти гонотрофических циклов, откладывая пять партий яиц и столько же раз насасываясь крови. При этом лишь очень небольшое количество крови расходуется на поддержание жизни, почти вся она используется для построения желтка яиц. В том случае, если самке не удаётся насосать полную порцию крови, развитие яиц вообще не наблюдается.
Несколько по-иному происходит образование яиц у бабочек. У непитающихся шелкопрядов (или коконопрядов) оогенез завершается до окрыления; у бражников часть яиц формируется также до окрыления, другая часть — после окрыления, за счёт дополнительного питания. У большинства дневных бабочек формирование яиц начинается после метаморфоза.
Ещё более своеобразна деятельность телотрофических овариол (см. рис. 137, Б), свойственных всем остальным насекомым, в частности равнокрылым, клопам и разноядным жукам. Отличие телетрофических овариол от политрофических состоит в том, что все трофоциты остаются в гермарии и связываются с проходящими в вителлярий ооцитами длинными желточными тяжами.
Обеспечивая процессы вителлогенеза необходимыми для синтеза желтка продуктами, трофоциты нередко остаются сгруппированными в цисты или сливаются в единый трофический синцитий. Все они участвуют также в формировании очередной партии яиц.
Очевидно, что столь интенсивные процессы оогенеза при участии всех трофоцитов овариолы требуют определённых гарантий реализации его продуктов, и, видимо, не случайно многие насекомые с телотрофическими овариолами приступают к формированию яиц, лишь обнаружив подходящий для их откладки субстрат. Не ограниченные гонотрофической гармонией, они с гораздо большей лёгкостью регулируют собственное половое состояние и имеют большие возможности для выбора необходимых для потомков условий.
Впрочем, различия овариол разных типов далеко не всегда выражены столь определённо. Реальные отношения характеризуются гораздо большим разнообразием вариантов, и здесь рассмотрены лишь морфофункциональные основы плодовитости и сексуального состояния самок.
Процессы вителлогенеза в овариолах всех типов обычно происходят лишь в последних фолликулах вителлярия, и общим источником вителлогенинов служит жировое тело. Эти специфичные для самок белки, проникая сквозь оболочку овариолы и фолликулярный эпителий, абсорбируются на поверхности яйцеклетки и затем переносятся внутрь посредством пиноцитоза. Иного рода белки, а также липиды и гликоген обычно транспортируются от трофоцитов, которые, выполнив свои функции, резорбируются ооцитом. После этого формирующееся яйцо облекается тончайшей желточной оболочкой и более толстым хорионом — скорлупой, секретируемой стенками фолликула. Обе эти оболочки (рис. 138) защищают яйцо, но не препятствуют газообмену и проникновению сперматозоидов. Последние проходят через специальные микропилярные отверстия, которые остаются в хорионе от тончайших плазматических нитей, связывающих ооцит с группой фолликулярных клеток.
Последующие процессы оогенеза уже не требуют присутствия яйца в вителлярии. Прорвав эпителиальную пробку у основания овариолы, оно выпадает (овулирует) в полость латерального яйцевода. Опорожнённый фолликул спадается, но ещё долгое время сохраняет следы прошедшей овуляции в виде распадающегося желтого тела (рис. 139). Впоследствии остатки фолликула почти полностью растворяются ферментами лизосом и не препятствуют овуляции яиц, развивающихся в следующих фолликулах. Освободившись от тормозящих влияний оостатического гормона, или антигонадотропина, они сразу приступают к вителлогенезу, в свою очередь притормаживая развитие более молодых ооцитов вителлярия.
Сформированное, но ещё диплоидное яйцо, проходя по латеральному и затем по непарному медиальному яйцеводам, минует проток семяприемника и оплодотворяется выходящим из него сперматозоидом. Продвигаясь далее, яйцо приступает к мейозу и облекается клейким секретом парных придаточных желез. Затем, обычно в метафазе первого деления, самка откладывает яйцо на избранный ею субстрат.
По сравнению с членистоногими других групп, обитающими в относительно постоянных условиях водной среды или в верхних горизонтах почвы, насекомые вынуждены приспосабливаться к более суровым условиям наземного существования. Вполне понятно, что вылупляющийся из яйца организм должен отличаться более высокой организацией и повышенной жизнеспособностью, что достигается благодаря совершенствованию эмбриогенеза и пополнению его новыми формообразовательными процессами. Это требует повышенных количеств желтка. Таким образом, развитие вителлярия (где накапливается желток) в овариолах насекомых можно рассматривать как приспособление к наземному обитанию. Во всяком случае, яичники членистоногих других групп лишены развитого вителлярия, а в их яйцах содержится значительно меньше желтка.
Среди факторов, регулирующих процессы вителлогенеза и созревания самок у насекомых, существенное значение имеют количество и качество корма. Приведённые примеры гонотрофических отношений у комаров можно дополнить примером зависимости развития яичников мух от содержания в пище белков и углеводов. При питании мух исключительно углеводной пищей сроки их жизни не сокращаются, но не образуется ни одного сформированного яйца. Включение в рацион белков сразу стимулирует развитие яичников через действие гормонов эндокринной системы.
Не менее показательно влияние плотности сосредоточения особей, повышение которой сокращает плодовитость мух дрозофил, но увеличивает её у пустынной саранчи. Если при этом имеются половозрелые самцы, то у саранчи стимулируется развитие яичников, по-видимому, благодаря феромонам. Впрочем, феромоны пчелиной матки, наоборот, предотвращают развитие яичников у рабочих особей.
Ещё более очевидно влияние температуры и влажности, оптимальным значениям которых соответствуют максимальная плодовитость, а также стимулирующее влияние на плодовитость введённого в половые пути самки сперматофора.
Особое значение в регуляции созревания самок и продуцировании ими яиц принадлежит сезонным изменениям фотопериода. Сигнализируя о неизбежном наступлении неблагоприятных условий осеннего времени, сокращение светового дня тормозит формирование яиц у многих насекомых умеренных и высоких широт, но служит положительным стимулом для обитателей засушливых тропиков. Все эти влияния опосредуются нервной и эндокринной системами, в частности нейросекреторными клетками головного мозга и ретроцеребрального комплекса. Некоторые примеры такого рода представлены схемами на рисунке 140.
Процессы оогенеза завершаются выделением полярных (направительных или редукционных) телец и формированием гаплоидного женского пронуклеуса.
По сравнению с процессами сперматогенеза оогенез менее продуктивен. В первом случае в результате мейоза образуются четыре сперматозоида, во втором — только одно яйцо и три дегенерирующих впоследствии полярных тельца.

