Распределительные системы и метаболизм
Кровеносная система.Редукция замкнутой кровеносной системы кольчатых червей оставила у их далёких потомков лишь незамкнутый спинной сосуд с сегментарно расположенными камерами, клапанами и остиями (рис. 88). Прилегая изнутри к тергитам брюшка, он всасывает гемолимфу и гонит её от слепого заднего конца вперёд, в лишённую клапанов и остий аорту. При этом полный цикл обращения гемолимфы в полости тела занимает не менее 6 мин.
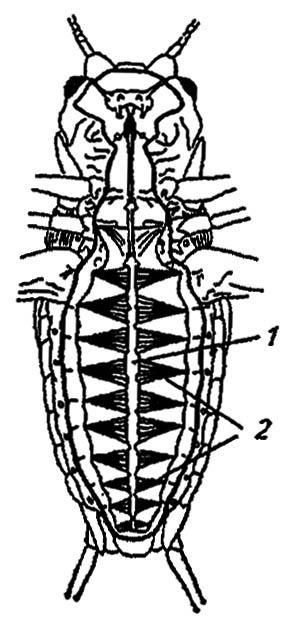
Рисунок 88. Спинной сосуд насекомых (по Gillot, 1980):
1 — спинной сосуд; 2 — крыловидные мышцы
Не участвуя в распределении кислорода, кровеносная система насекомых сохранила исходные функции переноса продуктов переваривания пищи от стенок кишечника к метаболически активным органам и поддержания гомеостаза, то есть постоянства внутренней среды организма, её защиты от агрессивных воздействий и агентов. Вместе с тем гемолимфа как полостная жидкость и кровь обеспечивает осмотическое и гидростатическое напряжения стенок тела, клеток и органов; являясь средой для протекающих в растворах биохимических реакций, она транспортирует и биологически активные соединения от мест их синтеза ко всем тканям.
Стенки спинного сосуда образованы кольцевыми мышцами миокарда с вплетёнными в них продольными и косыми волокнами и слоем соединительной ткани. Обычно сам миокард не способен к произвольным ритмическим и частым сокращениям; ему свойственны слабые волны перистальтики, зарождающиеся в его стенках миогенно. У некоторых насекомых спинной сосуд иннервируется латеральными нервами симпатической и сегментарными нервами центральной нервной системы. Поступающие по ним импульсы ускоряют ритм биений до 310 ударов в минуту, хотя обычно он не превышает 10–160 ударов. У комара Anopheles quadrimaculatus Mg. спинной сосуд вообще не иннервируется; его работа регулируется влиянием гормонов и физиологически активных веществ типа ацетилхолина.
Засасывая гемолимфу через остии, число которых варьирует от двенадцати до одной пары, спинной сосуд гонит её вперёд. Вслед за сокращением камер (фаза систолы) наступает расслабление миокарда (диастола), сменяемое фазой покоя (диастазис) между последовательными биениями. Движение гемолимфы обеспечивается также ритмичными колебаниями дорзальной и вентральной диафрагм, сокращениями крыловидных мышц и дополнительных пульсирующих органов (ампул и мембран) у основания крыльев, антенн и ног. Например, крыловые ампулы, имея собственный ритм сокращений, нагнетают гемолимфу в жилки костального края крыла. Затем она возвращается в тело по системам медиальных и анальных жилок.
Гемолимфа.
Составляя от 5 до 40 % массы тела, гемолимфа насекомых наряду с жидкой плазмой включает в себя форменные элементы — гемоциты. Обычно гемолимфа бесцветная, реже — жёлтая или зелёная, а у личинок комаров-звонцов, некоторых оводов и клопов-гладышей окрашена гемоглобином в красный цвет. Её реакция близка нейтральной (рН 6–7), но осмотическое давление заметно выше, чем у гемолимфы млекопитающих. Однако в качестве основных осмотически активных веществ здесь выступают аминокислоты и другие органические соединения, но не минеральные соли.
Особенно наглядны осмотические свойства гемолимфы у форм, заселяющих солёные и солоноватые воды. У личинок мух-береговушек Ephydra cinerea Flln. эти свойства остаются неизменными даже в крепком растворе соли. Излишние ионы выводятся мальпигиевыми сосудами и различными ректальными ампулами и железами, но в гипотонической среде они, наоборот, удерживают ионы в плазме.
В плазму входит вода с растворенными в ней газами (O2 и CO2) и диссоциированными неорганическими ионами (катионы: Na+, K+, Са2+, Mg2+ анионы: Cl-, HCO3-, H2PO4-), а также углеводы, многоатомные спирты, липиды, аминокислоты, пептиды, белки, пигменты и некоторые другие соединения, в частности относительно концентрированная мочевая кислота.
Из углеводов гемолимфы не менее 80 % приходится на долю трегалозы. Она синтезируется из двух молекул глюкозы в клетках жирового тела и расщепляется вновь ферментом трегалазой в метаболически активных органах. Далее отметим глицерин, синтезируемый здесь же из гликогена жирового тела при понижении температуры. Выполняя роль антифриза, он предотвращает замерзание гемолимфы зимующих насекомых. Например, у наездника Bracon cephi L. глицерин, составляя 25 % массы тела, снижает точку замерзания полостной жидкости до —17 °С. Среди липидов, присутствующих в гемолимфе обычно в виде липопротевдов, основная масса представлена сложными эфирами глицерина и высших жирных кислот. Последние служат источником энергии, и если трегалоза и гликоген расходуются уже в первые минуты полёта, то, например, жирные кислоты жирового тела у пустынной саранчи Schistocerca gregaria Forsk. обеспечивают непрерывный лёт в течение 5 ч.
Высокая концентрация аминокислот, в особенности глутамина и глутаминовой кислоты, определяется их ролью в осморегуляции. Вместе с тем они используются для синтеза белков и как резервы азотистых соединений для синтеза кутикулы (тирозина, триптофана). Поступая в гемолимфу из жирового тела, пищи и гидролизованных при метаморфозе белков тканей тела, они обычно сохраняются в виде пептидов. В гемолимфе присутствуют также белки, и их концентрация закономерно изменяется при линьках и развитии насекомого. Как субстрат морфогенетических процессов и источник энергии они участвуют в транспортировке углеводов и жиров в форме гликопротеидов и липопротеидов.
Следует отметить дегидрогеназы яблочной и изолимонной кислот — ферментов, способствующих осуществлению некоторых звеньев цикла Кребса (см. ниже) непосредственно в гемолимфе, а также специфичных для самок белков — вителлогенинов, необходимых для синтеза желтка в формирующихся яйцах. Имеющийся у многих насекомых белок — лизоцим служит одним из основных агентов защиты организма от болезнетворных патогенных бактерий.
Не менее разнообразны, чем клетки крови позвоночных животных, гемоциты насекомых. Гемоциты обычно оседают на стенках органов и лишь перед наступлением линьки и метаморфоза свободно плавают в плазме вместе с фрагментами тканей. Число их достигает 10 000–100 000 в 1 мм3 гемолимфы. Они формируются в специальных гемопоэтических органах, отмеченных в дорзальной диафрагме и близ спинного сосуда у сверчков, пилильщиков, чешуекрылых и мух.
Наиболее постоянны гемоциты трёх основных типов, тогда как шесть других типов встречаются спорадически и у немногих форм (рис. 89). По-видимому, все они берут начало от пролейкоцитов 5 — мелких округлых клеток с относительно крупным ядром и базофильной цитоплазмой. Испытывая митотические деления, пролейкоциты преобразуются в более крупные и весьма разнообразные по форме плазмоциты 6. Обладая гранулами и лизосомами в цитоплазме, они способны к фагоцитозу, как и гранулоциты, отличающиеся ацидофильной, заполненной гранулами цитоплазмой. Имеющиеся у двукрылых и перепончатокрылых сферулоциты 7 со сферическими включениями в ацидофильной цитоплазме, по-видимому, участвуют в транспорте гормонов, тогда как адипогемоциты 3 разносят жировые включения. Цистоциты 4, иногда именуемые коагулоцитами, обеспечивают свёртывание гемолимфы при ранениях тела насекомого. Наряду с ними изредка встречаются уплощённые подоциты 2, длинные червеобразные клетки 1 и крупные эноцитоиды 8 с 1–2 эксцентрично расположенными ядрами.
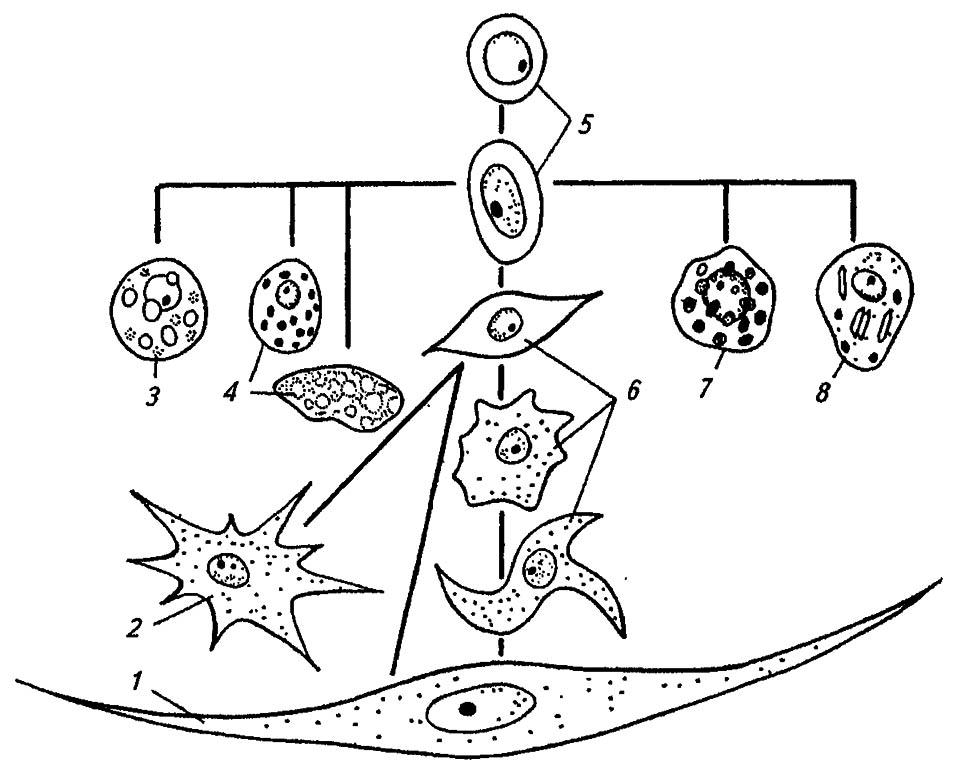
Рисунок 89. Различные типы гемоцитов насекомых и их взаимоотношения (по Тыщенко, 1976; Gillot, 1980)
Состав и относительную численность гемоцитов можно использовать для диагностики физиологического состояния организма.
Особое значение имеют барьерные, защитные свойства гемолимфы, проявляющиеся в фагоцитарной активности гемоцитов, в их агглютинации (склеивании) на поверхности ран, в бактерицидном действии лизоцима и наконец в инкапсуляции чужеродных включений и тел паразитов.
Представляют интерес сообщения о возможной иммунизации насекомых различными вакцинами. Правда, формирующийся иммунитет пассивен и скоротечен, так как ничего похожего на антитела у насекомых не обнаружено. Вероятно, в этих реакциях, описанных ещё в 1920 году С. И. Метальниковым, какую-то роль играют производные тирозина и недавно открытые простагландины.
Жировое тело.
Помимо гемоцитов и плазмы с кровеносной системой насекомых тесно связаны клетки жирового тела и атроциты — многоядерные перикардиальные клетки, перивисцеральные нефроциты, выполняющие экскреторные функции и регулирующие состав гемолимфы, а также эноциты. Последние в отличие от всех рассмотренных ранее компонентов имеют не мезодермальную, а эктодермальную природу и, секретируя липопротеиды и кутикулины, по-видимому, участвуют в метаболизме гормонов.
Продукты переваривания пищи, проникая в гемолимфу, обычно сразу же включаются в метаболизм. Почти во всех тканях и клетках вдет активная работа по синтезу специфических веществ, и если, например, в гиподерме образуется хитин, то в эпителии кишечника — ферменты. Однако средоточием всех основных процессов метаболизма и синтеза белков, жиров и углеводов служит жировое тело. В этом смысле оно аналог печени млекопитающих, а не метаболически пассивные отложения жира.
Клетки жирового тела так же, как и гемоциты, образуются из висцеральной мезодермы сомитов. Утрачивая изначальное распределение по сегментам, они вскоре рассредоточиваются по всей полости тела и связываются в пласты и дольки под покровами и близ кишечника. Сначала их трудно отличить от гемоцитов, но позже они сильно увеличиваются, приобретают обширные вакуоли и включения гликогена, жиров и белков, а их ядра становятся многолопастными. При наступлении метаморфоза эти крупные клетки распадаются, изливая содержимое своих тел в гемолимфу. У взрослых особей они восстанавливаются заново.
Основную массу жирового тела образуют клетки — трофоциты, способные синтезировать транспортные и резервные соединения. После 12-дневного голодания трофоциты личинок комара Aedes aegypti L. теряют все включения, а их вакуоли заполняются водой. Впоследствии, при потреблении личинками углеводов, они весьма быстро накапливают гликоген и некоторое количество жира, однако при питании растительными маслами трофоциты заполняются только жировыми включениями. Если голодающим личинкам предложить только белковую пищу, то в трофоцитах наряду с гранулами белка обнаруживаются жиры. Таким образом, липиды синтезируются при всех испытанных рационах, но в обычных условиях существования накопление гликогена и жира наблюдается только у личинок младших возрастов и лишь позднее резервные белки накапливаются у личинок старшего возраста.
В жировом теле насекомых имеются также уратные клетки, которые некоторые исследователи считают преобразованными трофоцитами. Они особенно многочисленны у форм, лишённых мальпигиевых сосудов, или в те периоды жизни, когда последние не функционируют. Накапливая в цитоплазме ураты и прочие конечные продукты, эти клетки избавляют от них другие, более активные ткани и таким образом дополняют функции обычных органов выделения.
У тараканов и некоторых других насекомых в жировом теле присутствуют мицетоциты — клетки, заполненные симбиотическими бактериями. Однако после скармливания насекомым антибиотиков бактерии исчезают и бывшие мицетоциты накапливают кристаллы мочевой кислоты.

