Выведение конечных продуктов метаболизма
Образовавшиеся при распаде пищи конечные продукты метаболизма либо выводятся через покровы тела и стенки трахей (CO2), либо абсорбируются в задней кишке (H2O), либо удаляются с остатками непереваренной пищи — экскрементами (мочевина, мочевая кислота, аммиак и др.).
При гидролизе нуклеиновых кислот образуются углеводы, фосфорная кислота и обогащённые азотом пуриновые (аденин, гуанин) или пиримидиновые (цитозин, тимин) основания. В свою очередь, пуриновые основания, подвергаясь окислению и дезаминированию, дают начало мочевой кислоте и её производным: аллантоину, аллантоиновой кислоте, мочевине и аммиаку, которые выводятся из организма. Пиримидиновые основания, хотя и способны преобразовываться в мочевину и аммиак, обычно вновь вовлекаются в метаболические процессы.
При гидролизе белков образуются аминокислоты и некоторые из них — чаще всего богатые азотом аргинин и гистидин — входят в состав экскрементов (в весьма малых количествах). Обычно они используются в синтезе пуриновых оснований, образуя наряду с ними мочевину. Таким образом, конечные продукты метаболизма азотсодержащих соединений формируются при окислении пуринов или синтезируются из аминокислот (рис. 100).
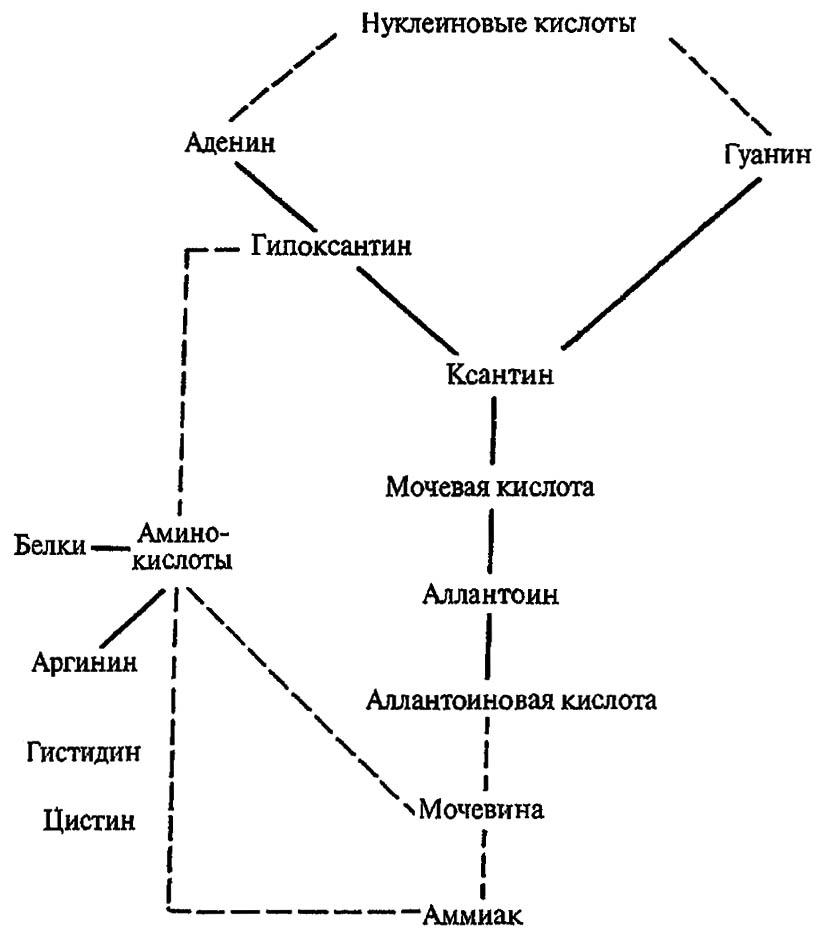
Рисунок 100. Конечные продукты обмена азотсодержащих соединений и их превращения у насекомых (по Gillot, 1980)
Большинство наземных насекомых выделяют азот в виде слаборастворимых и нетоксичных для организма мочевой кислоты, аллантоина и аллантоиновой кислоты. Они удаляются вместе с обезвоженными экскрементами; при этом возможные потери влаги сводятся к минимуму. Водорастворимые и токсичные даже в малых концентрациях мочевина и аммиак требуют для выведения очень больших количеств воды. Не случайно, что именно эти соединения являются конечными продуктами метаболизма у водных форм. Прежде чем поступить в заднюю кишку, в формирующиеся здесь экскременты, все эти метаболиты накапливаются в гемолимфе и извлекаются из неё специализированными органами выделения — мальпигиевыми сосудами.
Мальпигиевы сосуды представляют собой длинные и тонкие трубочки, впадающие в кишечник на уровне пилорического отдела (см. рис. 81). Вместе с задней кишкой они обеспечивают экскрецию азотсодержащих метаболитов и постоянство ионного баланса гемолимфы. Лишь у ногохвосток, некоторых двухвосток и тлей они не развиты.
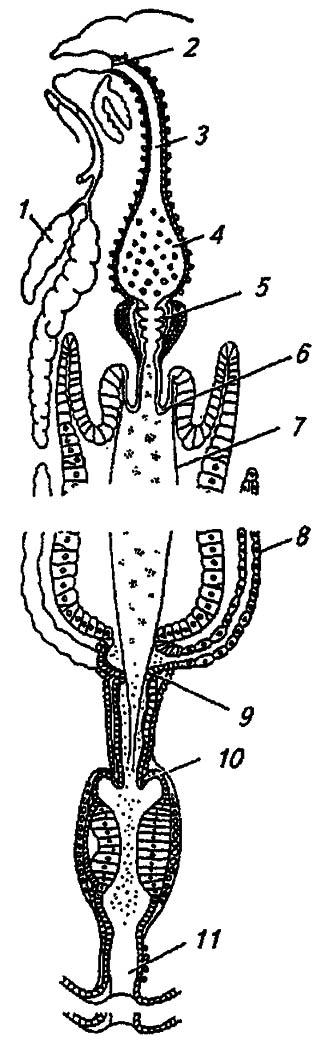
Рисунок 81. Схема кишечного тракта насекомых (по Шванвичу, 1949):
1 — слюнные железы; 2 — глотка; 3 — пищевод; 4 — зоб; 5 — провентрикулус; 6 — кардиальный клапан; 7 — перитрофическая мембрана; 8 — мальпигиев сосуд; 9, 10 — соответственно пилорический и ректальный клапаны; 11 — анус
Стенки сосудов образованы однослойным эпителием и мышечными волокнами. Оплетённые трахеями, но лишённые нервов, они способны только к миогенным червеобразным движениям. У щетино-хвосток, уховёрток и трипсов мальпигиевы сосуды не имеют мышц и пассивно колеблются в токах гемолимфы.
В простейшем случае, например у прямокрылых, мальпигиевы сосуды однообразны по всей длине и лишь насасывают плазму с содержащимися в ней экскретами (рис. 101). Далее эта «первичная моча» проникает в полость задней кишки и подвергается здесь реабсорбции. Все метаболически ценные вещества (H2O, Cl-, Na+, K+ и др.) возвращаются в гемолимфу, а экскреты выводятся из организма. Сравнительно малая эффективность работы таких сосудов компенсируется их громадным числом (до 250 и более).
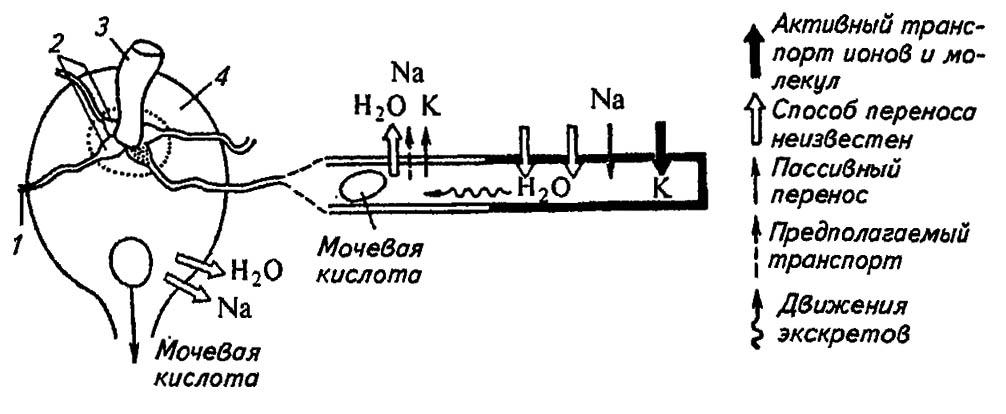
Рисунок 101. Строение и принципы работы мальпигиевых сосудов палочника (по Тыщенко, 1976):
1 — мальпигиевы сосуды; 2 — ампула; 3 — средняя кишка; 4 — задняя кишка
Сходным образом функционируют малочисленные (4–8) мальпигиевы сосуды некоторых жуков, однако их свободные концы врастают в стенку задней кишки. Высасывая из её полости воду, они энергично проводят первичную мочу, но не способны к её реабсорбции. У многих клопов происходит дифференциация отделов и эпителия сосудов и соответственно распределение функций по их длине. В дистальном отделе эпителиальные клетки несут плотный рабдориум и содействуют образованию первичной мочи. Переходя в проксимальный отдел, клетки которого снабжены рыхлым рабдориумом, она подвергается реабсорбции, и, таким образом, этот отдел принимает на себя функции задней кишки прямокрылых (рис. 102).
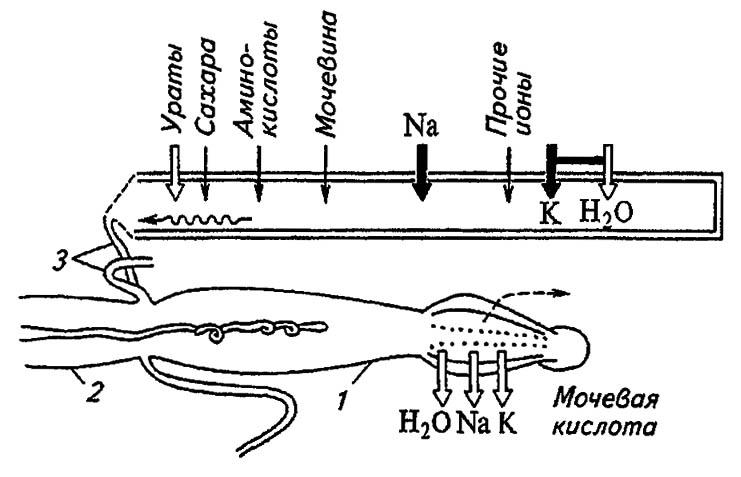
Рисунок 102. Строение и принципы работы мальпигиевых сосудов клопа Rhodnius prolixus St. (по Тыщенко, 1976):
1 — задняя кишка; 2 — средняя кишка; 3 — мальпигиевы сосуды
Ещё большей сложностью строения отличаются мальпигиевы сосуды двукрылых. Наряду с дистальным и проксимальным отделами в них выделяются промежуточный и медиальный отделы. В дистальном происходит насасывание мочевой кислоты и её солей, а также ионов Ca2+, тогда как в промежуточном и медиальном — воды. В проксимальком отделе реабсорбируются метаболически ценные продукты. У гусениц многих бабочек свойства сосудов, отмеченные у клопов и двукрылых, сочетаются с криптонефрией (рис. 103).
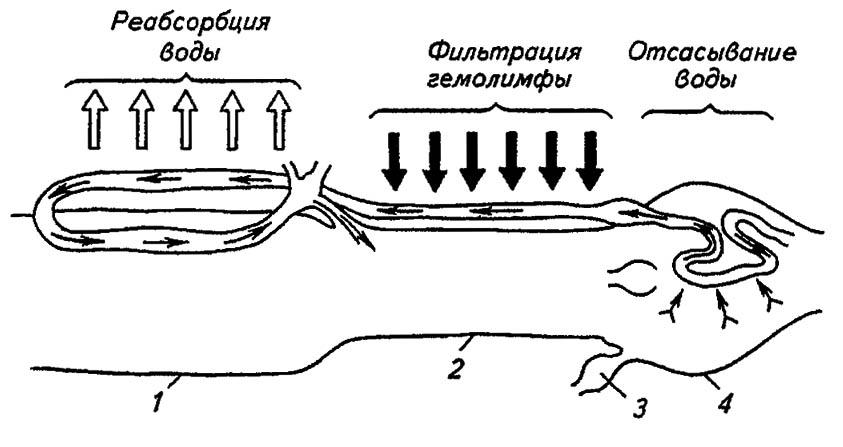
Рисунок 103. Строение и принципы работы мальпигиевых сосудов гусеницы бабочки Corcyra cephalonica (по Тыщенко, 1976):
1 — средняя кишка; 2 — тонкая кишка; 3 — ампула мальпигиева сосуда; 4 — прямая кишка
Заполняющая мальпигиевы сосуды жидкость изотонична гемолимфе, но отличается от неё по набору ионов. В частности, у палочника Carausius morosus Вr. ионы K+ преобладают внутри сосуда, а ионы Na+ — снаружи. Нарушение ионного баланса проявляется в разности потенциалов и возникновении электрохимического градиента.
Ионы K+ активно транспортируются внутрь и, по-видимому, переносят молекулы воды вопреки градиенту диффузии. Несколько по-иному работают мальпигиевы сосуды кровососущего клопа Rhodnius prolixus St. В них активно проникают ионы K+ и Na+, транспортирующие воду. Экскреты, поступающие в ихдистальные отделы в виде мочекислых солей натрия и калия, оказываются в слабощелочной среде (рН 7,2), но, продвигаясь проксимально, встречают слабокислую реакцию (рН 6,6) жидкости. В этих условиях Na+ и K+ освобождаются, а мочевая кислота кристаллизуется и выпадает в осадок (см. рис. 102).
Активность экскреции у Rhodnius prolixus St. существенно повышается (в 1 000 раз) под влиянием диуретического гормона, секретируемого в грудных ганглиях. Однако его выведение в гемолимфу происходит только при возбуждении рецепторов растяжения брюшка, что наблюдается всякий раз при насасывании крови. У саранчи Schistocerca gregaria Forsk. диуретический гормон стимулирует абсорбцию в мальпигиевых сосудах и тормозит реабсорбцию в ректальных железах задней кишки. У таракана Periplaneta americana L. наряду с диуретическим выделяется антидиуретический гормон.
Кроме мальпигиевых сосудов функции экскреции конечных продуктов метаболизма азота выполняют лабиальные железы Collembola, Thysanura и некоторых крылатых насекомых. У шелкопряда Hyalophora cecropia L. лабиальные шёлкоотделительные железы гусениц преобразуются в имагинальные органы, регулирующие водообмен и выделение экскретов. Продуцируемая придаточными половыми железами самцов некоторых тараканов мочевая кислота используется для покрытия сперматофоров и таким образом выводится из организма. Вместе с тем азотсодержащие метаболиты часто вообще не выводятся наружу, а, накапливаясь в уратных клетках жирового тела, в нефроцитах и в кутикуле, исключаются из процессов обмена веществ.
Согласованность и совершенство рассмотренных процессов метаболизма обеспечивают экономное расходование воды и энергетических субстратов, не допуская потерь сколько-нибудь ценных метаболитов. В этом отношении насекомые не уступают млекопитающим животным, несмотря на то что малые размеры тела определяют для них ряд ограничений. Однако ключевые пути метаболизма у тех и других принципиально сходны.

