5.7. Sperm competition
Multiple matings are common in many insect species. The occurrence of remating under natural conditions can be determined by observing the mating behavior of individual females or by dissection to establish the amount of ejaculate or the number of spermatophores present in the female’s sperm storage organs. Some of the best documentation of remating comes from studies of many Lepidoptera, in which part of each spermatophore persists in the bursa copulatrix of the female throughout her life (Fig. 5.6). These studies show that remating occurs, to some extent, in almost all species of Lepidoptera for which adequate field data are available.
The combination of internal fertilization, sperm storage, multiple mating by females, and the overlap within a female of ejaculates from different males leads to a phenomenon known as sperm competition. This occurs within the reproductive tract of the female at the time of oviposition when sperm from two or more males compete to fertilize the eggs. Both physiological and behavioral mechanisms determine the outcome of sperm competition. Thus, events inside the female’s reproductive tract, combined with various attributes of mating behavior, determine which sperm will succeed in reaching the eggs. It is important to realize that male reproductive fitness is measured in terms of the number of eggs fertilized or offspring fathered and not simply the number of copulations achieved, although these measures sometimes are correlated. Often there may be a trade-off between the number of copulations that a male can secure and the number of eggs that he will fertilize at each mating. A high copulation frequency is generally associated with low time or energy investment per copulation but also with low certainty of paternity. At the other extreme, males that exhibit substantial parental investment, such as feeding their mates (Boxes 5.1 & 5.2), and other adaptations that more directly increase certainty of paternity, will inseminate fewer females over a given period.
There are two main types of sexually selected adaptations in males that increase certainty of paternity. The first strategy involves mechanisms by which males can ensure that females use their sperm preferentially. Such sperm precedence is achieved usually by dis- placing the ejaculate of males that have mated previously with the female (Box 5.3). The second strategy is to reduce the effectiveness or occurrence of subsequent inseminations by other males. Various mechanisms appear to achieve this result, including mating plugs, use of male-derived secretions that “switch off ” female receptivity (Box 5.4), prolonged copulation (Fig. 5.8), guarding of females, and improved structures for gripping the female during copulation to prevent “takeover” by other males. A significant selective advantage would accrue to any male that could both achieve sperm precedence and prevent other males from successfully inseminating the female until his sperm had fertilized at least some of her eggs.
The factors that determine the outcome of sperm competition are not totally under male control. Female choice is a complicating influence, as shown in the above discussions on sexual selection and on morphology of genitalic structures. Female choice of sexual partners may be two-fold. First, there is good evidence that the females of many species choose among potential mating partners. For example, females of many mecopteran species mate selectively with males that provide food of a certain minimum size and quality (Box 5.1). In some insects, such as a few beetles and some moth and katydid species, females have been shown to prefer larger males as mating partners. Second, subsequent to copulation, the female might discriminate between partners as to which sperm will be used. One idea is that variation in the stimuli of the male genitalia induces the female to use one male’s sperm in preference to those of another, based upon an “internal courtship”. Differential sperm use is possible because females have control over sperm transport to storage, maintenance, and use at oviposition.
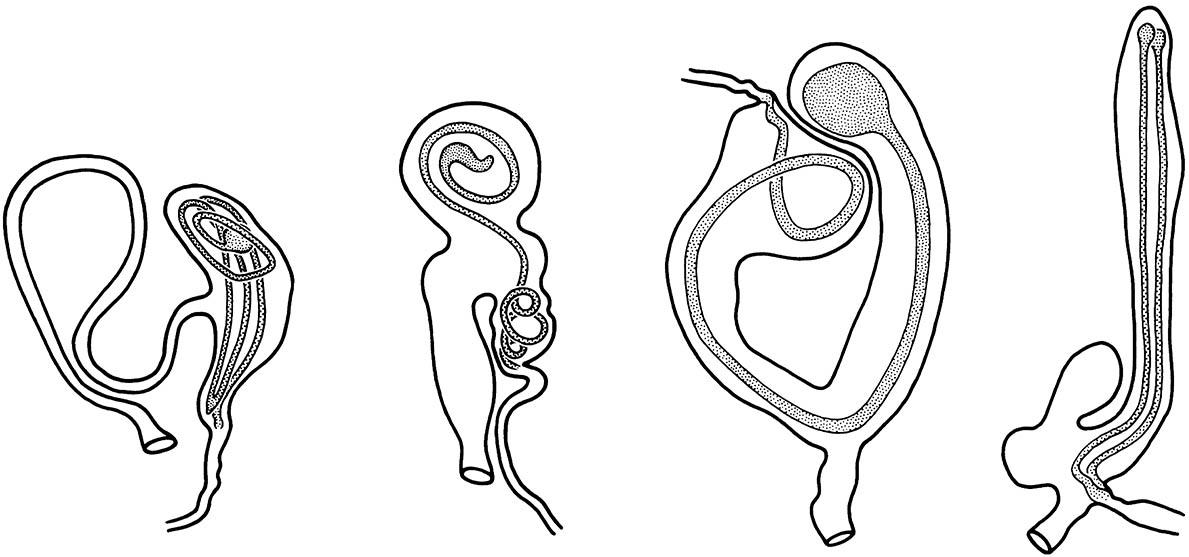
The sperm leave via the narrow end of each spermatophore, which has been deposited so that its opening lies opposite the “seminal duct” leading to the spermatheca (not drawn). The bursa on the far right contains two spermatophores, indicating that the female has remated. (After Williams 1941; Eberhard 1985)
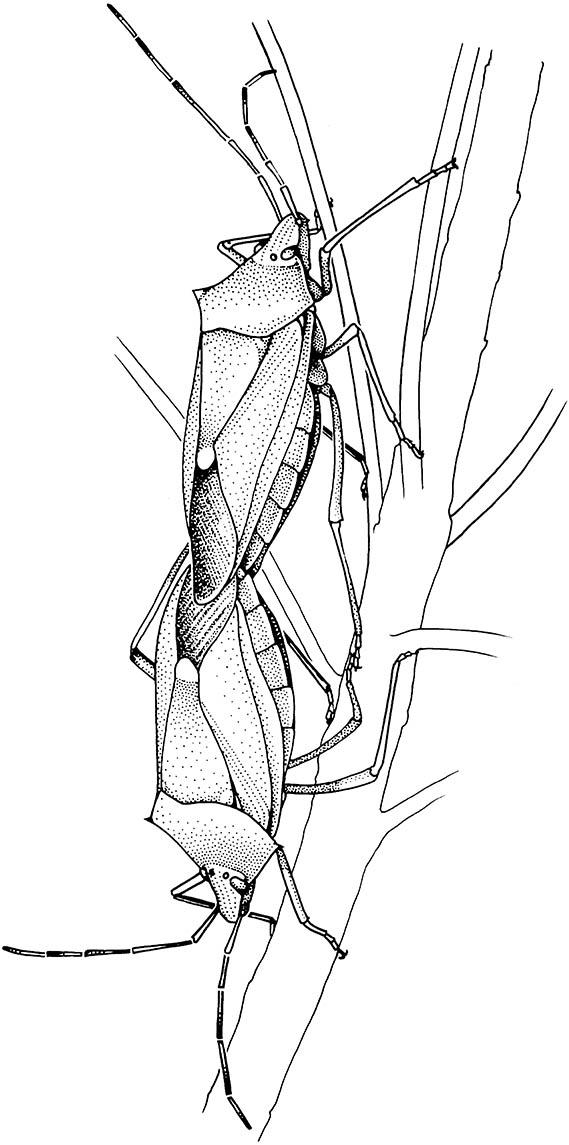
Many heteropteran bugs engage in prolonged copulation, which prevents other males from inseminating the female until either she becomes non-receptive to further males or she lays the eggs fertilized by the “guarding” male.

