Box 5.2. Nuptial feeding and other “gifts”
Feeding of the female by the male before, during, or after copulation has evolved independently in several different insect groups. From the female’s perspective, feeding takes one of three forms:
- receipt of nourishment from food collected, captured, or regurgitated by the male (Box 5.1); or
- obtaining nourishment from a glandular product (including the spermatophore) of the male; or
- by cannibalization of males during or after copulation.
From the male’s perspective, nuptial feeding may represent parental investment (provided that the male can be sure of his paternity), as it may increase the number or survival of the male’s offspring indirectly via nutritional benefits to the female. Alternatively, court- ship feeding may increase the male’s fertilization success by preventing the female from interfering with sperm transfer. These two hypotheses concerning the function of nuptial feeding are not necessarily mutually exclusive; their explanatory value appears to vary between insect groups and may depend, at least partly, on the nutritional status of the female at the time of mating. Studies of mating in Mecoptera, Orthoptera, and Mantodea exemplify the three nuptial feeding types seen in insects, and continuing research on these groups addresses the relative importance of the two main competing hypotheses that seek to explain the selective advantage of such feeding.
In some other insect orders, such as the Lepidoptera and Coleoptera, the female sometimes acquires metabolically essential substances or defensive chemicals from the male during copulation, but oral uptake by the female usually does not occur. The chemicals are transferred by the male with his ejaculate. Such nuptial gifts may function solely as a form of parental investment (as in puddling; see below) but may also be a form of mating effort (Box 14.3).
Puddling and sodium gifts in Lepidoptera
Male butterflies and moths frequently drink at pools of liquid, a behavior known as puddling . Anyone who has visited a tropical rainforest will have seen drinking clusters of perhaps hundreds of newly eclosed male butterflies, attracted particularly to urine, feces, and human sweat (see Plate 2.6, facing here). It has long been suggested that puddling — in which copious quantities of liquid are ingested orally and expelled anally — results in uptake of minerals, such as sodium, which are deficient in the larval (caterpillar) folivore diet. The sex bias in puddling occurs because the male uses the sodium obtained by puddling as a nuptial gift for his mate. In the moth Gluphisia septentrionis (Notodontidae) the sodium gift amounts to more than half of the puddler’s total body sodium and appears to be transferred to the female via his spermatophore (Smedley & Eisner 1996).
The female then apportions much of this sodium to her eggs, which contain several times more sodium than eggs sired by males that have been experimentally prevented from puddling. Such paternal investment in the offspring is of obvious advantage to them in supplying an ion important to body function.
In some other lepidopteran species, such “salted” gifts may function to increase the male’s reproductive fitness not only by improving the quality of his offspring but also by increasing the total number of eggs that he can fertilize, assuming that he remates. In the skipper butterfly, Thymelicus lineola (Hesperiidae), females usually mate only once and male-donated sodium appears essential for both their fecundity and longevity (Pivnick & McNeil 1987). These skipper males mate many times and can produce spermatophores without access to sodium from puddling but, after their first mating, they father fewer viable eggs compared with remating males that have been allowed to puddle. This raises the question of whether females, which should be selective in the choice of their sole partner, can discriminate between males based on their sodium load. If they can, then sexual selection via female choice also may have selected for male puddling.
In other studies, copulating male lepidopterans have been shown to donate a diversity of nutrients, including zinc, phosphorus, lipids, and amino acids, to their partners. Thus, paternal contribution of chemicals to offspring may be widespread within the Lepidoptera.
Mating in katydids (Orthoptera: Tettigoniidae)
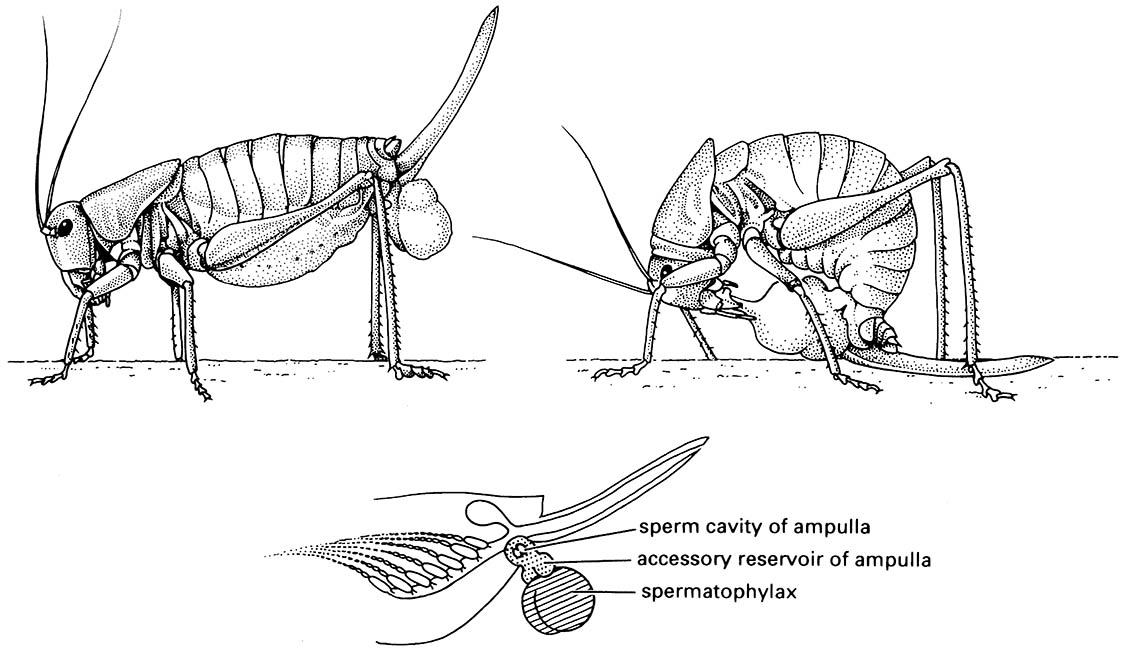
During copulation the males of many species of katydids transfer elaborate spermatophores, which are attached externally to the female’s genitalia (see Plate 3.1). Each spermatophore consists of a large, proteinaceous, sperm-free portion, the spermatophylax, which is eaten by the female after mating, and a sperm ampulla, eaten after the spermatophylax has been consumed and the sperm have been transferred to the female. The illustration (here) shows a recently mated female Mormon cricket, Anabrus simplex , with a spermatophore attached to her gonopore; in the illustration on the upper right, the female is consuming the spermatophylax of the spermatophore (after Gwynne 1981). The schematic illustration underneath depicts the posterior of a female Mormon cricket showing the two parts of the spermatophore: the spermatophylax (cross-hatched) and the sperm ampulla (stippled) (after Gwynne 1990). During consumption of the spermatophylax, sperm are transferred from the ampulla along with substances that “turn off ” female receptivity to further males. Insemination also stimulates oviposition by the female, thereby increasing the probability that the male supplying the spermatophore will fertilize the eggs.
There are two main hypotheses for the adaptive significance of this form of nuptial feeding. The spermatophylax may serve as a sperm-protection device by preventing the ampulla from being removed until after the complete ejaculate has been transferred. Alternatively, the spermatophylax may be a form of parental investment in which nutrients from the male increase the number or size of the eggs sired by that male. Of course, the spermatophylax may serve both of these purposes, and there is evidence from different species to support each hypothesis. Experimental alteration of the size of the spermatophylax has demonstrated that females take longer to eat larger ones, but in some katydid species the spermatophylax is larger than is needed to allow complete insemination and, in this case, the nutritional bonus to the female benefits the male’s offspring. The function of the spermatophylax apparently varies between genera, although phylogenetic analysis suggests that the ancestral condition within the Tettigoniidae was to possess a small spermatophylax that protected the ejaculate.
Cannibalistic mating in mantids (Mantodea)
The sex life of mantids is the subject of some controversy, partly as a consequence of behavioral observations made under unnatural conditions in the laboratory. For example, there are many reports of the male being eaten by the generally larger female before, during, or after mating. Males decapitated by females are even known to copulate more vigorously because of the loss of the suboesophageal ganglion that normally inhibits copulatory movements. Sexual cannibalism has been attributed to food deprivation in confinement but female mantids of at least some species may indeed eat their partners in the wild.
Courtship displays may be complex or absent, depending on species, but generally the female attracts the male via sex pheromones and visual cues. Typically, the male approaches the female cautiously, arresting movement if she turns her head towards him, and then he leaps onto her back from beyond her strike reach. Once mounted, he crouches to elude his partner’s grasp. Copulation usually lasts at least half an hour and may continue for several hours, during which sperm are transferred from the male to the female in a spermatophore. After mating, the male retreats hastily. If the male were in no danger of becoming the female’s meal, his distinctive behavior in the presence of the female would be inexplicable. Furthermore, suggestions of gains in reproductive fitness of the male via indirect nutritional benefits to his offspring are negated by the obvious unwillingness of the male to participate in the ultimate nuptial sacrifice — his own life!
Whereas there is no evidence yet for an increase in male reproductive success as a result of sexual cannibalism, females that obtain an extra meal by eating their mate may gain a selective advantage, especially if food is limiting. This hypothesis is supported by experiments with captive females of the Asian mantid Hierodula membranacea that were fed different quantities of food. The frequency of sexual cannibalism was higher for females of poorer nutritional condition and, among the females on the poorest diet, those that ate their mates produced significantly larger oothecae (egg packages) and hence more offspring. The cannibalized males would be making a parental investment only if their sperm fertilize the eggs that they have nourished. The crucial data on sperm competition in mantids are not available and so currently the advantages of this form of nuptial feeding are attributed entirely to the female.