12.2.3. Specialized hymenopterans: ants
Ants (Formicidae) form a well-defined, highly specialized group within the superfamily Vespoidea (Fig. 12.2). The morphology of a worker ant of Formica is illustrated in Box 12.2.
Colony and castes in ants
All ants are social and their species are polymorphic. There are two major female castes, the reproductive queen and the workers, usually with complete dimorphism between them. Many ants have monomorphic workers, but others have distinct subcastes called, according to their size, minor, media, or major workers. Although workers may form clearly different morphs, more often there is a gradient in size. Workers are never winged, but queens have wings that are shed after mating, as do males, which die after mating. Winged individuals are called alates. Polymorphism in ants is accompanied by polyethism, with the queen’s role restricted to oviposition, and the workers performing all other tasks. If workers are monomorphic, there may be temporal or age polyethism, with young workers undertaking internal nurse duties and older ones foraging outside the nest. If workers are polymorphic, the subcaste with the largest individuals, the major workers, usually has a defensive or soldier role.
The workers of certain ants, such as the fire ants (Solenopsis), have reduced ovaries and are irreversibly sterile. In others, workers have functional ovaries and may produce some or all of the male offspring by laying haploid (unfertilized) eggs. In some species, when the queen is removed, the colony continues to produce gynes from fertilized eggs previously laid by the queen, and males from eggs laid by workers. The inhibition by the queen of her daughter workers is quite striking in the African weaver ant, Oecophylla longinoda. A mature colony of up to half a million workers, distributed amongst as many as 17 nests, is prevented from reproduction completely by a single queen. Workers, however, do produce male offspring in nests that lie outside the influence (or territory) of the queen. Queens prevent the production of reproductive eggs by workers, but may allow the laying of specialized trophic eggs that are fed to the queen and/or larvae. By this means the queen not only prevents any reproductive com- petition, but directs much of the protein in the colony towards her own offspring.
Caste differentiation is largely trophogenic (diet- determined), involving biased allocation of volume and quality of food given to the larvae. A high-protein diet promotes differentiation of gyne/queen and a less rich, more dilute diet leads to differentiation of workers. The queen generally inhibits the development of gynes indirectly by modifying the feeding behavior of workers towards female larvae, which have the potential to differentiate as either gynes or workers. In Myrmica, large, slowly developing larvae will become gynes, so stimulation of rapid development and early metamorphosis of small larvae, or food deprivation and irritating of large larvae by biting to accelerate development, both induce differentiation as workers. When queen influence wanes, either through the increased size of the colony, or because the inhibitory pheromone is impeded in its circulation throughout the colony, gynes are produced at some distance from the queen. There is also a role for JH in caste differentiation. JH tends to induce queen development during egg and larval stages, and induces production of major workers from already differentiated workers.
According to a seasonal cycle, ant gynes mature to winged reproductives, or alates, and remain in the nest in a sexually inactive state until external conditions are suitable for departing the nest. At the appropriate time they make their nuptial flight, mate, and attempt to found a new colony.
Nesting in ants
The subterranean soil nests of Myrmica and the mounds of plant debris of Formica are typical temperate ant nests. Colonies are founded when a mated queen sheds her wings and overwinters, sealed into a newly dug nest that she will never leave. In spring, the queen lays some eggs and feeds the hatched larvae by stomodeal or oral trophallaxis, i.e. regurgitation of liquid food from her internal food reserves. Colonies develop slowly whilst worker numbers build up, and a nest may be many years old before alates are produced.
Colony foundation by more than one queen, known as pleometrosis, appears to be fairly widespread, and the digging of the initial nest may be shared, as in the honeypot ant Myrmecocystus mimicus. In this species and others, multi-queen nests may persist as polygynous colonies, but monogyny commonly arises through dominance of a single queen, usually following rearing of the first brood of workers. Polygynous nests often are associated with opportunistic use of ephemeral resources, or persistent but patchy resources.
The woven nests of Oecophylla species are well- known, complex structures (Fig. 12.7). These African and Asian/Australian weaver ants have extended territories that workers continually explore for any leaf that can be bent. A remarkable collaborative construction effort follows, in which leaves are manipulated into a tent-shape by linear ranks of workers, often involving “living chains” of ants that bridge wide gaps between the leaf edges. Another group of workers take larvae from existing nests and carry them held delicately between their mandibles to the construction site. There, larvae are induced to produce silk threads from their well-developed silk glands and a nest is woven linking the framework of leaves.
Living plant tissues provide a location for nests of ants such as Pseudomyrmex ferrugineus, which nests in the expanded thorns of the Central American bull’s- horn acacia trees (Fig. 11.10a). In such mutualisms involving plant defense, plants benefit by deterrence of phytophagous animals by the ants, as discussed in section 11.4.1.
Foraging efficiency of ants can be very high. A typical mature colony of European red ants (Formica polyctena) is estimated to harvest about 1 kg of arthropod food per day. The legionary, or army, and driver ants are popularly known for their voracious predatory activities. These ants, which predominantly belong to the subfamilies Ecitoninae and Dorylinae, alternate cyclically between sedentary (statary) and migratory or nomadic phases. In the latter phase, a nightly bivouac is formed, which often is no more than an exposed cluster of the entire colony. Each morning, the millions-strong colony moves in toto, bearing the larvae. The advancing edge of this massive group raids and forages on a wide range of terrestrial arthropods, and group predation allows even large prey items to be overcome. After some two weeks of nomadism, a statary period commences, during which the queen lays 100,000–300,000 eggs in a statary bivouac. This is more sheltered than a typical overnight bivouac, perhaps within an old ants’ nest, or beneath a log. In the three weeks before the eggs hatch, larvae of the previous oviposition complete their development to emerge as new workers, thus stimulating the next migratory period.
Not all ants are predatory. Some ants harvest grain and seeds (myrmecochory; section 11.3.2) and others, including the extraordinary honeypot ants, feed almost exclusively on insect-produced honeydew, including that of scale insects tended inside nests (section 11.4.1). Workers of honeypot ants return to the nest with crops filled with honeydew, which is fed by oral trophallaxis to selected workers called repletes. The abdomen of repletes are so distensible that they become virtually immobile “honey pots” (Fig. 2.4), which act as food reserves for all in the nest.

The superfamily Apoidea includes the Sphecidae sensu stricto, the Crabronidae (formerly part of a broader Sphecidae), the Ampulicidae (not shown), and all bees, here treated as one family, the Apidae, with several subfamilies (e.g. Apinae, Colletinae, Halictinae; not all solitary groups are shown) of uncertain relationships. Traditionally, bees have been classified in several families, a ranking that is unjustified phylogenetically. Probable relationships within non-social aculeate wasps (e.g. Ampulicidae, Pompilidae, and Rhopalosomatidae) and bees are not depicted. (Adapted from several sources including Gauld & Bolton 1988; Alexander 1992; Brothers 1999; B.N. Danforth, pers. comm.)

(After CSIRO 1970; Hölldobler 1984)
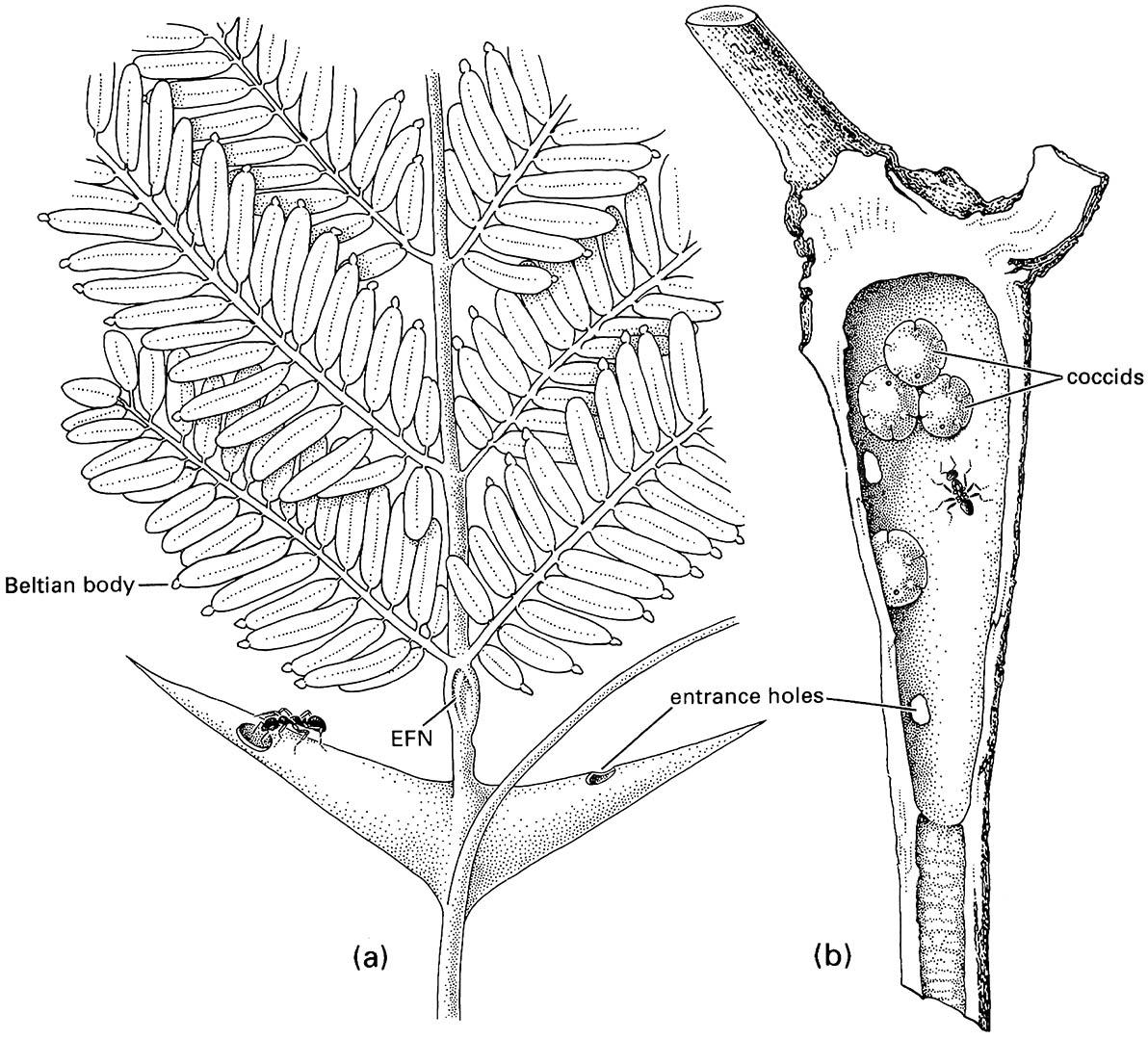
(a) a neotropical bull’s-horn acacia, Acacia sphaerocephala (Fabaceae), with hollow thorns, food bodies, and extrafloral nectaries (EFNs) that are used by the resident Pseudomyrmex ants; (b) a hollow swollen internode of Kibara (Monimiaceae) with scale insects of Myzolecanium kibarae (Hemiptera: Coccidae) that excrete honeydew that is eaten by the resident ants of Anonychomyrma scrutator. ((a) After Wheeler 1910; (b) after Beccari 1877)
The arthrodial membrane between tergal plates is depicted to the right in its unfolded and folded conditions. (After Hadley 1986; Devitt 1989)

