12.2.2. Specialized eusocial hymenopterans: wasps and bees
The highly eusocial hymenopterans comprise the ants (family Formicidae) and some wasps, notably Vespinae, and many bees, including most Apinae (Figs. 12.2 & 12.3). Bees are derived from sphecid wasps and differ from wasps in anatomy, physiology, and behavior in association with their dietary specialization. Most bees provision their larvae with nectar and pollen rather than animal material. Morphological adaptations of bees associated with pollen collection include plumose (branched) hairs, and usually a widened hind basitarsus adorned with hairs in the form of a brush (scopa) or a fringe surrounding a concavity (the corbicula, or pollen basket) (Fig. 12.4). Pollen collected on the body hairs is groomed by the legs and transferred to the mouthparts, scopae, or corbiculae. The diagnostic features and the biology of all hymenopterans are dealt with in Box 12.2, which includes an illustration of the morphology of a worker vespine wasp and a worker ant.
Colony and castes in eusocial wasps and bees
The female castes are dimorphic, differing markedly in their appearance. Generally, the queen is larger than any worker, as in vespines such as the European wasps (Vespula vulgaris and V. germanica), and honey bees (Apis spp.). The typical eusocial wasp queen has a differentially (allometrically) enlarged gaster (abdomen). In worker wasps the bursa copulatrix is small, preventing mating, even though in the absence of a queen their ovaries will develop.
In the vespine wasps, the colony-founding queen, or gyne, produces only workers in the first brood.
Immediately after these are hatched, the queen wasp ceases to forage and devotes herself exclusively to reproduction. As the colony matures, subsequent broods include increasing proportions of males, and finally gynes are produced late in the season from larger cells than those from which workers are produced.
The tasks of vespine workers include:
- distribution of protein-rich food to larvae and carbo-hydrate-rich food to adult wasps;
- cleaning cells and disposal of dead larvae;
- ventilation and air-conditioning of the nest by wing-fanning;
- nest defense by guarding entrances;
- foraging outside for water, sugary liquids, and insect prey;
- construction, extension, and repair of the cells and inner and outer nest walls with wood pulp, which is masticated to produce paper.
Each worker is capable of carrying out any of these tasks, but often there is an age polyethism: newly emerged workers tend to remain in the nest engaged in construction and food distribution. A middle-aged foraging period follows, which may be partitioned into wood-pulp collection, predation, and fluid-gathering phases. In old age, guarding duties dominate. As newly recruited workers are produced continuously, the age structure allows flexibility in performing the range of tasks required by an active colony. There are seasonal variations, with foraging occupying much of the time of the colony in the founding period, with fewer resources — or a lesser proportion of workers’ time — devoted to these activities in the mature colony. Male eggs are laid in increasing numbers as the season progresses, perhaps by queens, or by workers on whom the influence of the queen has waned.
The biology of the honey bee, Apis mellifera, is extremely well studied because of the economic significance of honey and the relative ease of observing honey-bee behavior (Box 12.1). Workers differ from queens in being smaller, possessing wax glands, having a pollen-collecting apparatus comprising pollen combs and a corbicula on each hind leg, in having a barbed sting that cannot be retracted after use, and in some other features associated with the tasks that workers perform. The queen’s sting is scarcely barbed and is retractible and reusable, allowing repeated assaults on pretenders to the queen’s position. Queens have a shorter proboscis than workers and lack several glands.
Honey-bee workers are more or less monomorphic, but exhibit polyethism. Thus, young workers tend to be “hive bees”, engaged in within-hive activities, such as nursing larvae and cleaning cells, and older workers are foraging “field bees”. Seasonal changes are evident, such as the 8—9-month longevity of winter bees, com- pared with the 4—6-week longevity of summer workers. Juvenile hormone (JH) is involved in these behavioral changes, with levels of JH rising from winter to spring, and also in the change from hive bees to field bees. Honey-bee worker activities correlate with seasons, notably in the energy expenditure involved in thermoregulation of the hive.
Caste differentiation in honey bees, as in eusocial hymenopterans generally, is largely trophogenic, i.e. determined by the quantity and quality of the larval diet. In species that provision each cell with enough food to allow the egg to develop to the pupa and adult without further replenishment, differences in the food quantity and quality provided to each cell determine how the larva will develop. In honey bees, although cells are constructed according to the type of caste that is to develop within them, the caste is determined neither by the egg laid by the queen, nor by the cell itself, but by food supplied by workers to the developing larva (Fig. 12.5). The type of cell guides the queen as to whether to lay fertilized or unfertilized eggs, and identifies to the worker which type of rearing (principally food) to be supplied to the occupant. Food given to future queens is known as “royal jelly” and differs from worker food in having a high sugar content and being composed predominantly of mandibular gland products, namely pantothenic acid and biopterin. Eggs and larvae up to three days old can differentiate into queens or workers according to upbringing. However, by the third day a potential queen has been fed royal jelly at up to 10 times the rate of less-rich food supplied to a future worker. At this stage, if a future queen is transferred to a worker cell for further development, she will become an intercaste, a worker-like queen. The opposite trans- fer, of a three-day-old larva reared as a worker into a queen cell, gives rise to a queen-like worker, still retaining the pollen baskets, barbed stings, and mandibles of a worker. After four days of appropriate feeding, the castes are fully differentiated and transfers between cell types result in either retention of the early determined outcome or failure to develop.
Trophogenic effects cannot always be separated from endocrine effects, as nutritional status is linked to corpora allata activity. It is clear that JH levels correlate with polymorphic caste differentiation in eusocial insects. However, there seems to be much specific and temporal variation in JH titers and no common pattern of control is yet evident.
The queen maintains control over the workers’ reproduction principally through pheromones. The mandibular glands of queens produce a compound identified as (E)-9-oxodec-2-enoic acid (9-ODA), but the intact queen inhibits worker ovarian development more effectively than this active compound. A second pheromone has been found in the gaster of the queen, and this, together with a second component of the mandibular gland, effectively inhibits ovarian development. Queen recognition by the rest of the colony involves a pheromone disseminated by attendant workers that contact the queen and then move about the colony as messenger bees. Also, as the queen moves around on the comb whilst ovipositing into the cells, she leaves a trail of footprint pheromone. Production of queens takes place in cells that are distant from the effects of the queen’s pheromone control, as occurs when nests become very large. Should the queen die, the volatile pheromone signal dissipates rapidly, and the workers become aware of the absence. Honey bees have very strongly developed chemical communication, with specific pheromones associated with mating, alarm, and orientation as well as colony recognition and regulation. Physical threats are rare, and are used only by young gynes towards workers.
Males, termed drones, are produced throughout the life of the honey-bee colony, either by the queen or perhaps by workers with developed ovaries. Males contribute little to the colony, living only to mate: their genitalia are ripped out after copulation and they die.
Nest construction in eusocial wasps
The founding of a new colony of eusocial vespid wasps takes place in spring, following the emergence of an overwintering queen. After her departure from the natal colony the previous fall, the new queen mates, but her ovarioles remain undeveloped during the temperature-induced winter quiescence or facultative diapause. As spring temperatures rise, queens leave hibernation and feed on nectar or sap, and the ovarioles grow. The resting site, which may be shared by several overwintering queens, is not a prospective site for foundation of the new colony. Each queen scouts individually for a suitable cavity and fighting may occur if sites are scarce.
Nest construction begins with the use of the mandibles to scrape wood fibers from sound or, more rarely, rotten wood. The wasp returns to the nest site using visual cues, carrying the wood pulp masticated with water and saliva in the mandibles. This pulpy paper is applied to the underside of a selected support at the top of the cavity. From this initial buttress, the pulp is formed into a descending pillar, upon which is suspended ultimately the embryonic colony of 20–40 cells (Fig. 12.6). The first two cells, rounded in cross-section, are attached and then an umbrella-like envelope is formed over the cells. The envelope is elevated by about the width of the queen’s body above the cells, allowing the queen to rest there, curled around the pillar. The developing colony grows by the addition of further cells, now hexagonal in cross-section and wider at the open end, and by either extension of the envelope or construction of a new one. The queen forages only for building material at the start of nest construction. As the larvae develop from the first cells, both liquid and insect prey are sought to nourish the developing larvae, although wood pulp continues to be collected for further cell construction. This first embryonic phase of the life of the colony ceases as the first workers emerge.
As the colony grows, further pillars are added, providing support to more lateral areas where brood-filled cells are aligned in combs (series of adjoining cells aligned in parallel rows). The early cells and envelopes become overgrown, and their materials may be reused in later construction. In a subterranean nest, the occupants may have to excavate soil and even small stones to allow colony expansion, resulting in a mature nest (as in Fig. 12.6), which may contain as many as 12,000 cells. The colony has some independence from external temperature, as thoracic heating through wing beating and larval feeding can raise temperature, and high temperature can be lowered by directional fanning or by evaporation of liquid applied to the pupal cells.
At the end of the season, males and gynes (potential queens) are produced and are fed with larval saliva and prey brought into the nest by workers. As the old queen fails and dies, and gynes emerge from the nest, the colony declines rapidly and the nest is destroyed as workers fight and larvae are neglected. Potential queens and males mate away from the nest, and the mated female seeks a suitable overwintering site.
Nesting in honey bees
In honey bees, new colonies are initiated if the old one becomes too crowded. When a bee colony becomes too large and the population density too high, a founder queen, accompanied by a swarm of workers, seeks a new nest site. Because workers cannot survive long on the honey reserves carried in their stomachs, a suitable site must be found quickly. Scouts may have started the search several days before formation of the swarm. If a suitable cavity is found, the scout returns to the cluster and communicates the direction and quality of the site by a dance (Box 12.1). Optimally, a new site should be beyond the foraging territory of the old nest, but not so distant that energy is expended in long-distance flight. Bees from temperate areas select enclosed nest sites in cavities of about 40 liters in volume, whereas more tropical bees choose smaller cavities or nest outside.
Following consensus over the nest site, workers start building a nest using wax. Wax is unique to social bees and is produced by workers that metabolize honey in fat cells located close to the wax glands. These modified epidermal cells lie beneath wax mirrors (overlapping plates) ventrally on the fourth to seventh abdominal segments. Flakes of wax are extruded beneath each wax mirror and protrude slightly from each segment of a worker that is actively producing wax. Wax is quite malleable at the ambient nest temperature of 35°C, and when mixed with saliva can be manipulated for cell construction. At nest foundation, workers already may have wax protruding from the abdominal wax glands. They start to construct combs of back-to-back hexagonal cells in a parallel series, or comb. Combs are separated from one another by pillars and bridges of wax. A thick cell base of wax is extended into a thin-walled cell of remarkably constant dimensions, despite a series of workers being involved in construction. In contrast to other social insects such as the vespids described above, cells do not hang downwards but are angled at about 13° above the horizontal, thereby preventing loss of honey. The precise orientation of the cells and comb derives from the bees’ ability to detect gravity through the proprioceptor hair plates at the base of their necks. Although removal of the hair plates prevents cell construction, worker bees could construct serviceable cells under conditions of weightlessness in space.
Unlike most other bees, honey bees do not chew up and reuse wax: once a cell is constructed it is permanently part of the nest, and cells are reused after the brood has emerged or the food contents have been used. Cell sizes vary, with small cells used to rear workers, and larger ones for drones (Fig. 12.5). Later in the life of the nest, elongate conical cells in which queens are reared are constructed at the bottom and sides of the nest. The brood develops and pollen is stored in lower and more central cells, whereas honey is stored in upper and peripheral cells. Workers form honey primarily from nectar taken from flowers, but also from secretions from extrafloral nectaries, or insect-produced honeydew. Workers carry nectar to the hive in honey stomachs, from which it may be fed directly to the brood and to other adults. However, most often it is converted to honey by enzymatic digestion of the sugars to simpler forms and reduction of the water content by evaporation before storage in wax-sealed cells until required to feed adults or larvae. It has been calculated that in 66,000 bee-hours of labor, 1 kg of beeswax can be formed into 77,000 cells, which can support the weight of 22 kg of honey. An average colony requires about 60–80 kg of honey per annum.
Unlike wasps, honey bees do not hibernate with the arrival of the lower temperatures of temperate winter. Colonies remain active through the winter, but foraging is curtailed and no brood is reared. Stored honey provides an energy source for activity and heat generation within the nest. As outside temperatures drop, the workers cluster together, heads inwards, forming an inactive layer of bees on the outside, and warmer, more active, feeding bees on the inside. Despite the prodigious stores of honey and pollen, a long or extremely cold winter may kill many bees.
Beehives are artificial constructions that resemble feral honey-bee nests in some dimensions, notably the distance between the combs. When given wooden frames separated by an invariable natural spacing interval of 9.6 mm honey bees construct their combs within the frame without formation of the internal waxen bridges needed to separate the combs of a feral nest. This width between combs is approximately the space required for bees to move unimpeded on both combs. The ability to remove frames allows the apiculturalist (beekeeper) to examine and remove the honey, and replace the frames in the hive. The ease of construction allows the building of several ranks of boxes. The hives can be transported to suitable locations with- out damaging the combs. Although the apiculture industry has developed through commercial production of honey, lack of native pollinators in monocultural agricultural systems has led to increasing reliance on the mobility of hive bees to ensure the pollination of crops as diverse as canola, nuts, soybeans, fruits, clover, alfalfa, and other fodder crops. In the USA alone, in 1998 some 2,500,000 bee colonies were rented for pollination purposes and the value to US agriculture attributable to honey-bee pollination was about US$15 billion in 2000. Yield losses of over 90% of fruit, seed, and nut crops would occur without honey-bee pollination. The role of the many species of eusocial native bees is little recognized, but may be important in areas of natural vegetation.

The superfamily Apoidea includes the Sphecidae sensu stricto, the Crabronidae (formerly part of a broader Sphecidae), the Ampulicidae (not shown), and all bees, here treated as one family, the Apidae, with several subfamilies (e.g. Apinae, Colletinae, Halictinae; not all solitary groups are shown) of uncertain relationships. Traditionally, bees have been classified in several families, a ranking that is unjustified phylogenetically. Probable relationships within non-social aculeate wasps (e.g. Ampulicidae, Pompilidae, and Rhopalosomatidae) and bees are not depicted. (Adapted from several sources including Gauld & Bolton 1988; Alexander 1992; Brothers 1999; B.N. Danforth, pers. comm.)
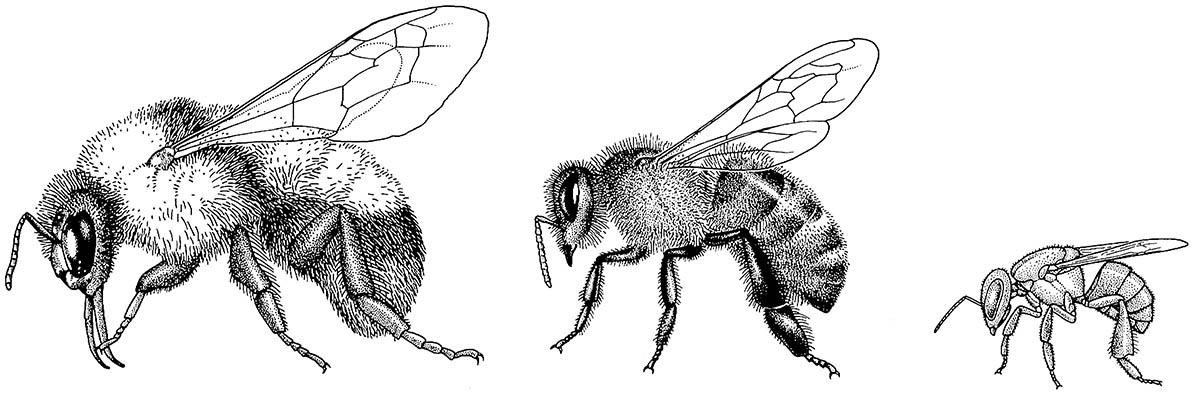
(After various sources, especially Michener 1974)
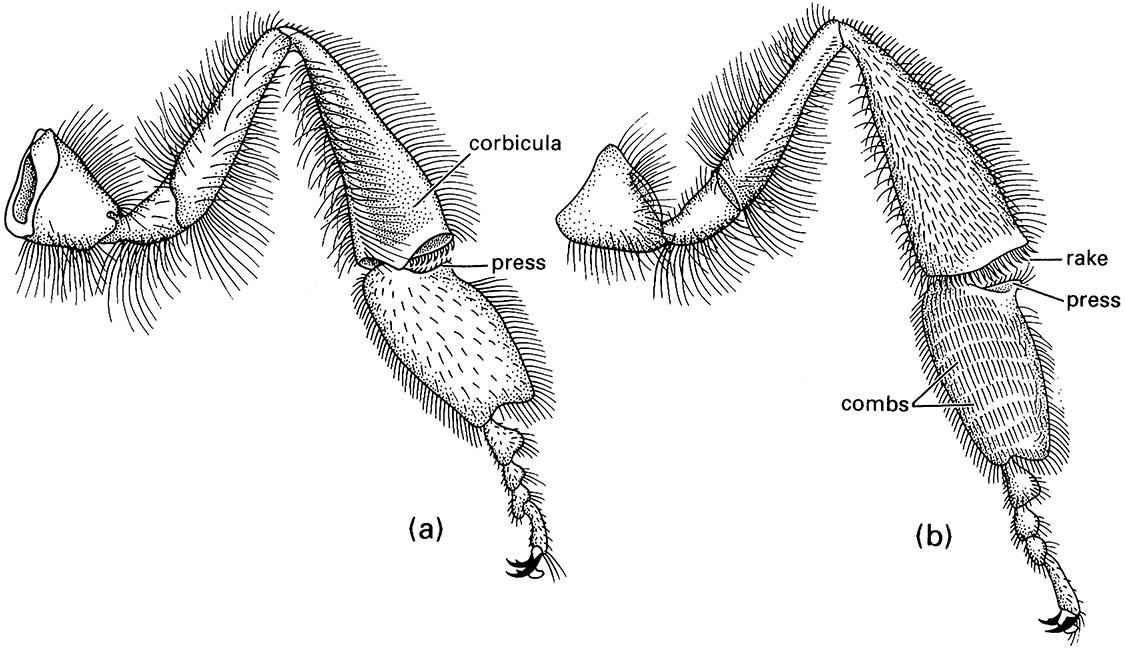
(a) outer surface showing corbicula, or pollen basket (consisting of a depression fringed by stiff setae), and the press that pushes the pollen into the basket; (b) the inner surface with the combs and rakes that manipulate pollen into the press prior to packing. (After Snodgrass 1956; Winston 1987)
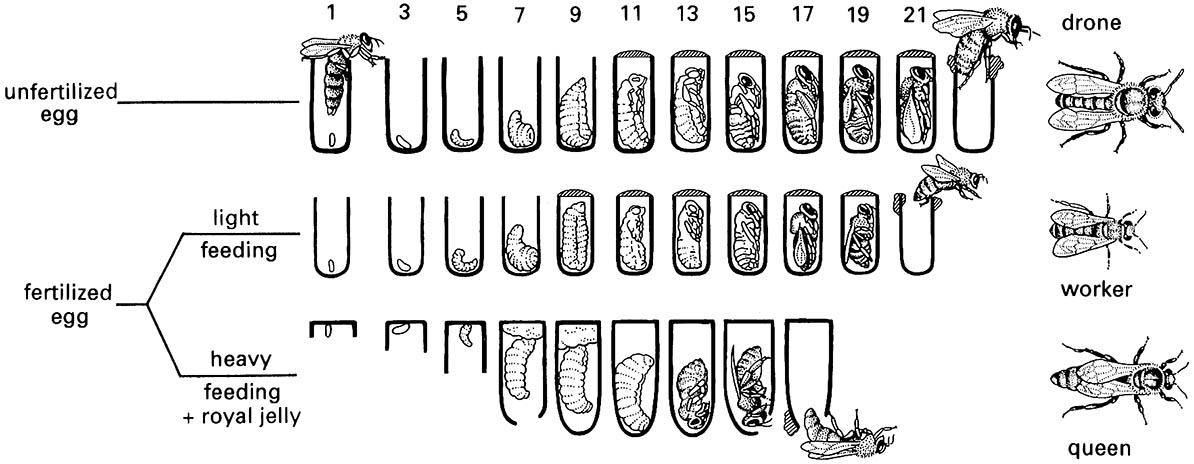
(After Winston 1987)
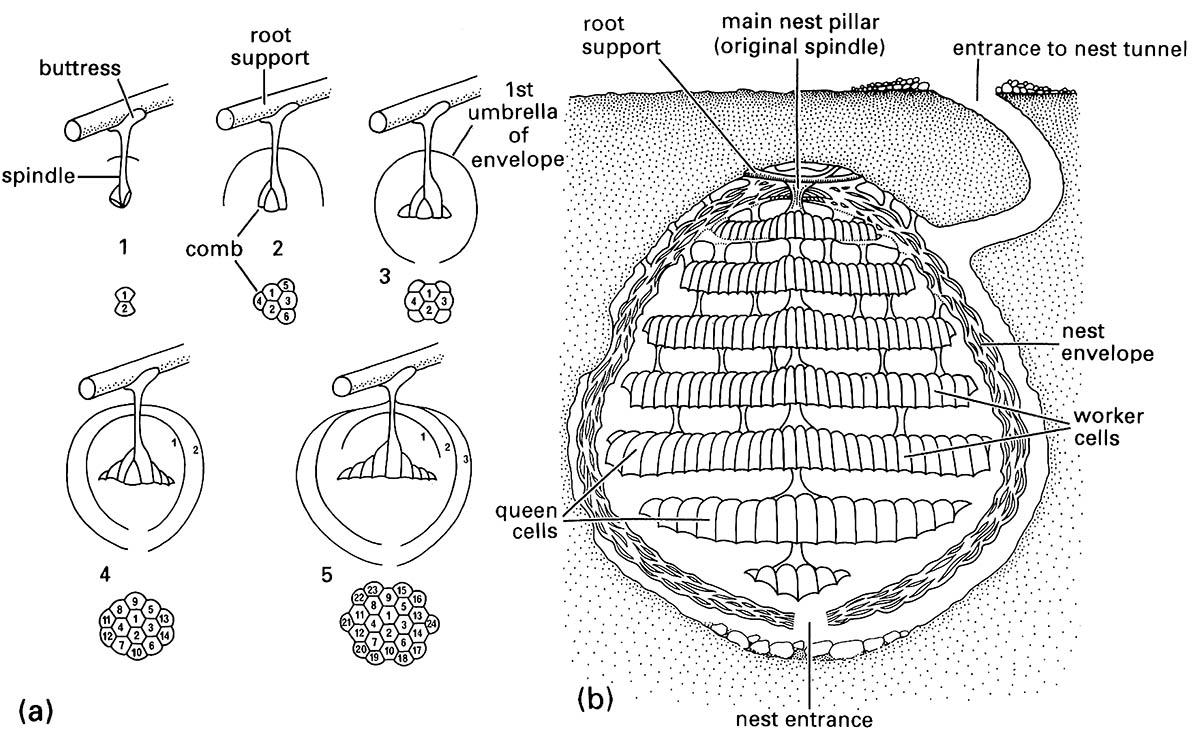
(a) initial stages (1-5) of nest construction by the queen (the embryonic phase of the colony’s life); (b) a mature nest. (After Spradbery 1973)

