Box 12.1. The dance language of bees
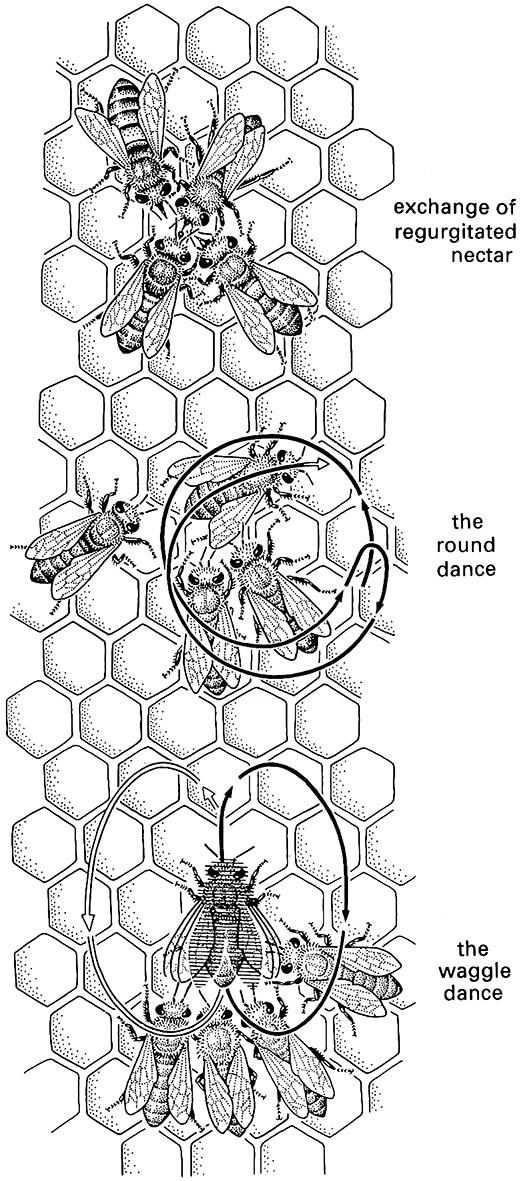
Honey bees have impressive communication abilities. Their ability to communicate forage sites to their nest- mates first was recognized when a marked worker provided with an artificial food source was allowed to return to its hive, and then prevented from returning to the food. The rapid appearance at the food source of other workers indicated that information concerning the resource had been transferred within the hive. Subsequent observations using a glass-fronted hive showed that foragers often performed a dance on return to the nest. Other workers followed the dancer, made antennal contact and tasted regurgitated food, as depicted here in the upper illustration (after Frisch 1967). Olfactory communication alone could be dis- counted by experimental manipulation of food sources, and the importance of dancing became recognized. Variations within different dances allow communication and recruitment of workers to close or distant food sources, and to food versus prospective nest sites. The purpose and messages associated with three dances — the round, waggle, and dorsoventral abdominal vibrating (DVAV) — have become well understood.
Nearby food is communicated by a simple round dance involving the incoming worker exchanging nectar and making tight circles, with frequent reversals, for a few seconds to a few minutes, as shown in the central illustration (after Frisch 1967). The quality of nectar or pollen from the source is communicated by the vigor of the dance. Although no directionality is conveyed, 89% of 174 workers contacted by the dancer during a round dance were able to find the novel food source within five minutes, probably by flying in ever increasing circles until the local source is found.
More distant food sources are identified by a waggle dance, which involves abdomen-shaking during a figure-of-eight circuit, shown in the lower illustration (after Frisch 1967), as well as food sharing. Informative characteristics of the dance include the length of the straight part (measured by the number of comb cells traversed), the dance tempo (number of dances per unit time), the duration of waggling and noise production (buzzing) during the straight-line section, and the orientation of the straight run relative to gravity. Messages conveyed are the energy required to get to the source (rather than absolute distance), quality of the forage, and direction relative to the sun’s position (Box 4.4). This interpretation of the significance and information content of the waggle dance was challenged by some experimentalists, who ascribed food-site location entirely to odors particular to the site and borne by the dancer. The claim mainly centered on the protracted time taken for the worker observers of the dance to locate a specific site. The duration matches more the time expected for a bee to take to locate an odor plume and subsequently zig-zag up the plume (Fig. 4.7), compared with direct flight from bearings provided in the dance. Following some well-designed studies, it is now evident that food finding is as effective and efficient when the experimental source is placed upwind as when it is placed downwind. Furthermore, although experienced workers can locate food by odor, the waggle dance serves to communicate information to naïve workers that allows them to head in the correct general direction. Close to the food source, specific odor does appear to be significant, and the final stages of orientation may be the slow part of location (particularly in experimental set-ups, with non-authentic food sources stationed beside human observers).
The function of the vibration dance (DVAV) differs from the round and waggle dances in regulating the daily and seasonal foraging patterns in relation to fluctuating food supply. Workers vibrate their bodies, particularly their abdomens, in a dorsoventral plane, usually whilst in contact with another bee. Vibration dances peak at times of the day and season when the colony needs to be primed for increased foraging, and these dances act to recruit workers into the waggle-dance area. Vibration dancing with queen contact appears to lessen the inhibitory capacity of the queen, and is used during the period when queen rearing is taking place. Cessation of this kind of vibration dancing may result in the queen departing with a swarm, or in the mating flight of new queens.
Communication of a suitable site for a new nest differs somewhat from communication of a food source. The returning scout dances without any nectar or pollen and the dance lasts for 15–30 minutes rather than the 1–2 minutes’ duration of the forager’s dance. At first, several scouts returning from various prospective new sites will all dance, with differences in tempo, angle, and duration that indicate the different directions and quality of the sites, as in a waggle dance. More scouts then fly out to prospect and some sites are rejected. Gradually, a consensus is attained, as shown by one dance that indicates the agreed site.
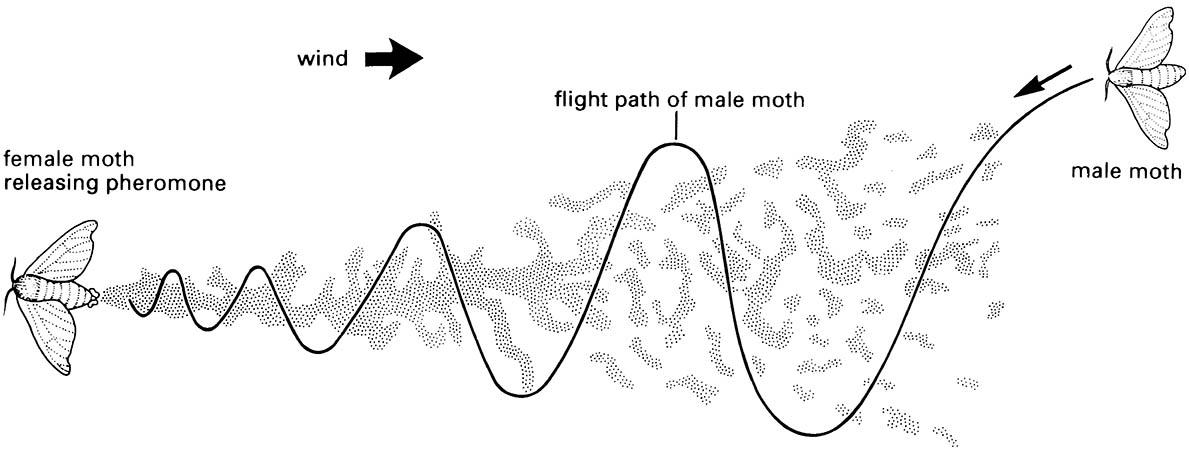
The pheromone trail forms a somewhat discontinuous plume because of turbulence, intermittent release, and other factors. (After Haynes & Birch 1985)