8.3. Were the first insects aquatic or terrestrial?
Arthropods evolved in the sea. This belief is based on evidence from the variety of forms preserved in Cambrian-age marine-derived deposits, such as the Burgess Shale in Canada and the Qiongzhusi Formation at Chengjiang in southern China. It is commonly believed that insects evolved after their hexapod ancestors become terrestrial, rather than insects making the ancestral transition from the ocean to land via estuaries and freshwater. The main evidence in support of a terrestrial origin for the Insecta derives from the fact that all extant non-pterygote insects (the apterygotes) and the other hexapods (Diplura, Collembola, and Protura) are terrestrial. That is, all the early-branching taxa in the hexapod phylogenetic tree (Fig. 7.2) live on land and there is no evidence from fossils (either by their possessing aquatic features or from details of preservation site) to suggest that the ancestors of these groups were not terrestrial (although they may have been associated with the margins of aquatic habitats). In contrast, the juveniles of five pterygote orders (Ephemeroptera, Odonata, Plecoptera, Megaloptera, and Trichoptera) live almost exclusively in freshwater. Given the positions of the Ephemeroptera and Odonata in Fig. 7.2, the ancestral condition for the protopterygotes probably involved immature development in freshwater.
Another line of evidence against an aquatic origin for the earliest insects is the difficulty in envisaging how a tracheal system could have evolved in water. In an aerial environment, simple invagination of external respiratory surfaces and subsequent internal elaboration could have given rise to a tracheal system (as shown in Fig. 8.3a) that later served as a preadaptation for tracheal gas exchange in the gills of aquatic insects (as shown in Fig. 8.3b). Thus, gill-like structures could assume an efficient oxygen uptake function (more than just diffusion across the cuticle) but only after the evolution of tracheae in a terrestrial ancestor.
There is no single explanation as to why virtually all insects with aquatic immature development have retained an aerial adult stage. Certainly, retention has occurred independently in several lineages (such as a number of times within both the Coleoptera and Diptera). The suggestion that a flighted adult is a predator-avoidance mechanism seems unlikely as predation could be avoided by a motile aquatic adult, as with so many crustaceans. It is conceivable that an aerial stage is retained to facilitate mating — perhaps there are mechanical disadvantages to underwater copulation in insects, or perhaps mate recognition systems may not function in water, especially if they are pheromonal or auditory.
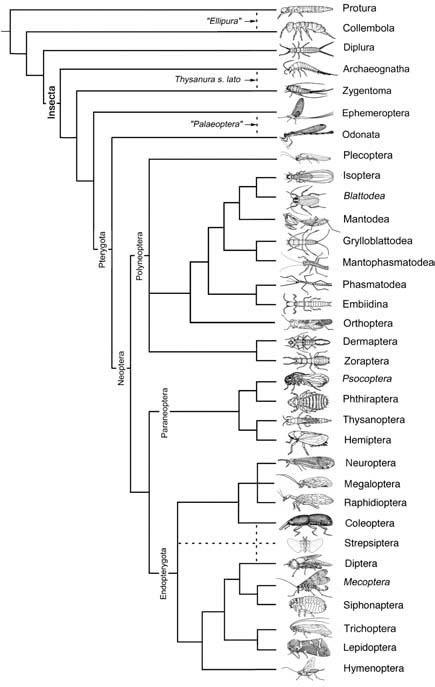
Broken lines indicate uncertain relationships. Thysanura sensu lato refers to Thysanura in the broad sense. (Data from several sources)
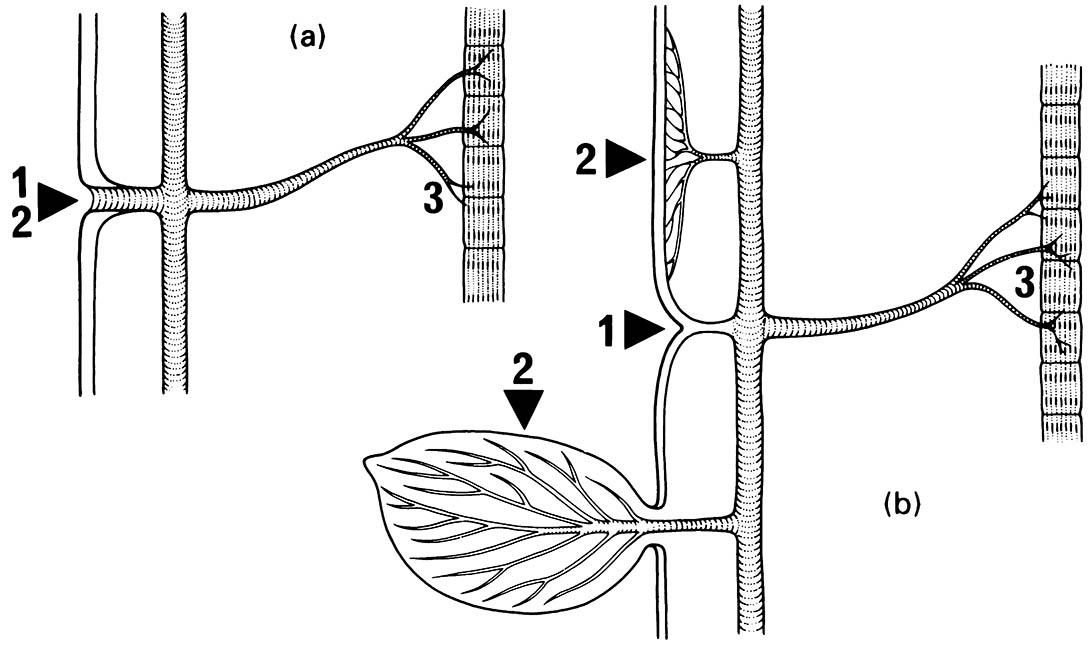
(a) Oxygen uptake through invagination. (b) Invagination closed, with tracheal gas exchange through gill. 1, indicates point of invagination of the tracheal system; 2, indicates point for oxygen uptake; 3, indicates poi nt for oxygen delivery, such as muscles. (After Pritchard et al. 1993)

