Факторы динамики численности популяций
Насекомые приобретают значение вредителей сельскохозяйственных культур только в том случае, если их численность превосходит экономические пороги вредоносности, поскольку отдельное, даже самое прожорливое насекомое не в состоянии нанести сколько-нибудь существенный вред урожаю. Поэтому планирование защитных мероприятий и соответствующие научные исследования направлены на сокращение до этих порогов числа особей в популяциях.
Динамика численности популяций насекомых проявляется либо в сезонном изменении их численности на протяжении года, либо на протяжении ряда лет, приобретая при этом благодаря исключительной энергии размножения многих видов характер закономерно чередующихся популяционных волн (рис. 193).
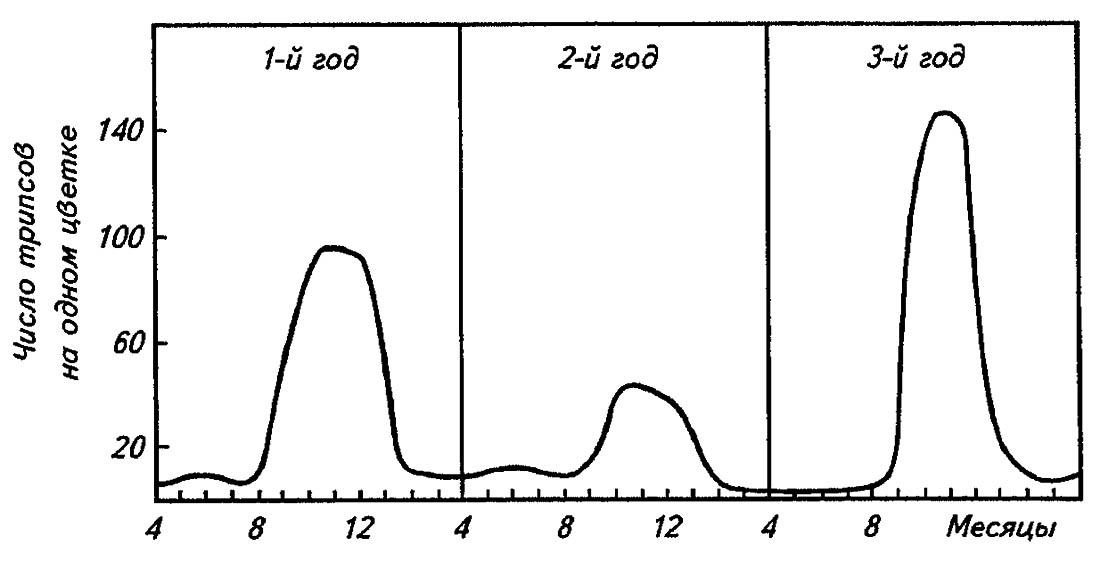
Рисунок 193. Популяционные волны динамики численности трипса (по Дажо, 1975)
Изменения численности популяций насекомых в реальной природной обстановке определяются факторами разного рода: климатом (абиотические факторы), существованием и активностью других организмов (внутривидовые и межвидовые отношения) и в значительной степени последствиями хозяйственной деятельности человека (антропогенные факторы).
Чрезвычайное многообразие этих факторов и сложность опосредуемых ими влияний обусловливают существование различных концепций роли отдельных факторов в динамике численности популяций насекомых.
В монофакториальных концепциях значение ведущего регулятора численности популяций придаётся одному важнейшему фактору, тогда как сторонники полифакториальной концепции рассматривают регуляцию численности как результат совместного действия многих факторов.
Относительная простота концепций первого типа содействовала разработке математических моделей, например описанной выше модели хищник — жертва. Однако нестабильность таких систем из-за элиминации либо хищника, либо жертвы ограничивает использование моделей экспериментальными ситуациями, требующими введения дополнительных факторов стабилизации численности. В природных условиях факторами такого рода могут стать миграции, не учитываемые в модели. Общий недостаток всех монофакториальных концепций, связывающих колебания численности либо с активностью хищника и паразита, либо с устойчивостью кормового растения, либо с наличием доступных убежищ и тому подобного, заключается в том, что эти концепции не учитывают неизбежность регулярности колебаний численности популяций близ некоторого уровня стабильности. Природные взаимосвязи намного разнообразнее наблюдаемых в экспериментальных ситуациях.
Для расчленения такого многообразного и ёмкого понятия, как сопротивление среды, на ряд факторов, ограничивающих экспоненциальный рост популяции, рассмотрим данные таблицы 4, рассчитанные для малинно-земляничного долгоносика (Anthonomus rubi Hbst.).
Очевидно, что здесь роль болезней как фактора снижения численности популяций долгоносика существенно уступает роли хищников и паразитов, что стадия личинки наиболее уязвима, а имаго наиболее защищена.
Такая дифференциация факторов позволяет понять их реальное значение в существовании и воспроизводстве популяций, и такого рода таблицы выживания наряду с представленными выше становятся важнейшим инструментом познания механизмов динамики численности.
Современные подходы к дифференциации факторов численности популяций исходят из трудов Говарда и Фиске (1911), противопоставивших факторы факультативные, постоянно усиливающие своё действие по мере возрастания численности, катастрофические, не зависящие от численности популяций, и стабильные, действие которых постоянно и не зависимо от численности.
Впоследствии факультативные факторы были сведены к факторам, зависящим от плотности популяций, факторы катастрофические — к не зависящим от плотности, а значение стабильных факторов существенно ограничилось. Существуют две противоположные точки зрения о роли факторов разных категорий в регуляции численности популяций. Полагая, что уровень численности определяется факторами, не зависящими от плотности популяций, сторонники одной точки зрения ссылаются на редкость сочетания условий, необходимых для постоянного роста популяций. Примеры массовых размножений насекомых, по их мнению, — редкие исключения из правила и выражают специфические свойства немногих видов. Численность популяций подавляющего большинства видов лимитируется краткостью сроков, когда сочетания условий обеспечивают рост популяции. При этом основными, лимитирующими численность факторами можно считать ограниченность ресурсов, их относительную недоступность при слабом развитии миграционных и поисковых способностей, а также скоротечность периода, когда рождаемость преобладает над смертностью, а скорость роста популяций положительна. Однако случайность колебаний численности в ответ на изменение условий, не связанных с плотностью популяций, рано или поздно приведёт популяции к нижним пределам численности и вымиранию.
Придерживаясь другой точки зрения, которая отдаёт предпочтение факторам, зависящим от плотности популяций, исследователи — приверженцы противоположного направления — сформулировали концепцию автоматического регулирования численности популяций. До недавнего времени поиски критериев для оценки регулирующей роли указанных факторов ограничивались лишь плотностью популяций, которая сокращается, если превышается некоторый средний её уровень, или, наоборот, увеличивается, если этот уровень остаётся недостигнутым. Используя кривые, выражающие динамику численности популяций на разных её уровнях и регрессионный анализ зависимости плотности популяций предыдущего и последующего поколений, можно по значению коэффициента регрессии судить о роли регулирующих факторов. Если коэффициент равен 1, то популяции не испытывают влияний факторов, зависящих от их плотности, и отклонения от этой величины выявляют степень участия таких факторов в динамике численности.
Современное состояние концепции автоматического регулирования численности популяций базируется на сочетании двух принципиально различных явлений: модификаций, или случайных колебаний численности, и регуляций, действующих по принципу кибернетической обратной связи и нивелирующих эти колебания.
В соответствии с этим противопоставляются модифицирующие (не зависящие от плотности популяций) и регулирующие (зависящие от плотности популяций) экологические факторы, причём первые из них воздействуют на организмы либо непосредственно, либо через изменения других компонентов биоценоза. По существу,
они представляют собой различные абиотические факторы. Регулирующие факторы связывают с существованием и активностью других живых организмов (биотические факторы), поскольку лишь живые существа способны реагировать на плотность своей собственной популяции и популяций других видов по принципу отрицательной обратной связи (рис. 194).
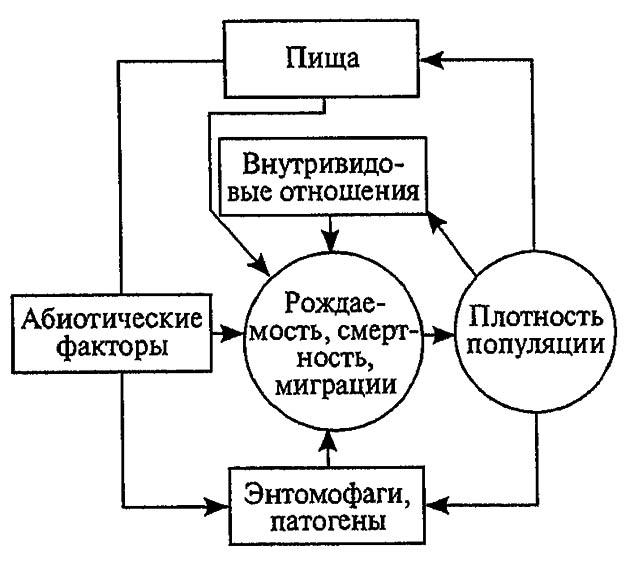
Рисунок 194. Факторы динамики численности популяции с позиций концепции автоматического регулирования (по Викторову, 1976)
Если воздействия модифицирующих факторов приводят лишь к преобразованиям (модификациям) колебаний численности, не устраняя их по существу, то регулирующие факторы, выравнивая случайные отклонения, стабилизируют (регулируют) численность на определённом уровне. Однако на разных уровнях численности популяции регулирующие факторы принципиально различны, например многоядные хищники и паразиты, способные при изменении численности жертв сокращать или усиливать свою активность (функциональная реакция), оказывают действие на сравнительно низких её уровнях (рис. 195).
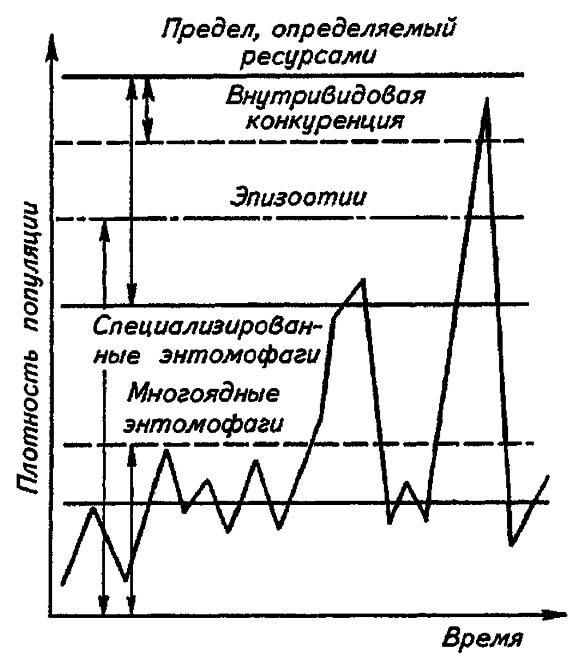
Рисунок 195. Включение разных регулирующих механизмов на разных уровнях численности популяций (по Викторову, 1976)
Специализированные энтомофаги, характеризующиеся численной реакцией на состояние популяций жертв, оказывают регулирующее действие в более широком диапазоне. При достижении популяцией жертвы ещё более высокой численности создаются условия для распространения болезней, и, наконец, предельный фактор регуляции согласно излагаемой концепции — внутривидовая конкуренция, ведущая к исчерпанию доступных ресурсов и развитию стрессовых реакций.
Таким образом, в данной схеме учитывается существование многозвенной буферной системы регуляции на основе биотических факторов среды, степень влияния которых зависит от плотности популяций.
В настоящее время концепция автоматического регулирования численности подверглась серьёзным критическим замечаниям.
В частности, при анализе динамики численности популяций массовых и редких видов насекомых вообще не было выявлено многозвенной буферной системы регуляции; характер отмечаемых флуктуаций и процесс их регуляции у тех и других видов оказались тождественными, а регистрируемые различия касались лишь уровней численности, на которых отмечались данные флуктуации.
Не отрицая значения плотности популяции как важной характеристики её состояния, следует признать, что изменения иных параметров не менее существенны. С состоянием и динамикой популяций связывают такие параметры, как генофонд популяции, распределение по стациям, структура скрещиваний, и многие другие. Иными словами, объективное описание популяции невозможно, если влияния экологических факторов учитывают только посредством изменений её плотности. Более того, оптимальная регуляция численности некоторых видов насекомых выявляется в ситуациях, когда в ответ на изменения численности под влиянием регулирующих факторов сохраняется возможность миграции некоторой части популяции за пределы ареала. При этом в каждом конкретном случае соотношения факторов меняются, и нередко сам процесс регуляции осуществляется за счёт миграций на новые территории, где особи либо находят для себя благоприятные условия, либо гибнут. Роль миграции в регуляции численности, особенно редких видов, изучена ещё крайне слабо, но, например, у дубовой зелёной листовёртки (Tortrix viridana L.) — массового вида мигранты появляются лишь на очень высоком уровне численности и при этом становятся устойчивыми к действию плотностно-зависимых факторов.
В существующих схемах регуляции численности популяций почти не учтена их генетическая разнокачественность, и часто явно недооценивается роль внутреннего фактора, то есть особенностей, присущих виду, популяции и членам популяции. При этом считают, что относительная роль «внутреннего фактора» в сравнении с ролью внешних факторов невелика, и потому необъективно оценивают реальность.
Проведение анализа эколого-генетической структуры популяций необходимо, но при этом следует учитывать возможность изменения свойств популяций при изменении их численности.
Представление об учтённых нами факторах динамики численности популяций насекомых и взаимодействиях между ними даёт схема, приведённая на рисунке 196. Выделение в данной схеме областей, соответствующих сопротивлению среды (K) и присущей популяции способности к росту (r), выражает взаимодействие основных факторов, а противопоставление рождаемости и смертности (соответственно иммиграции и эмиграции) — возможность действия этих факторов через изменения либо рождаемости (иммиграции), либо смертности (эмиграции). Введение в схему нового компонента — структуры популяции — соответствует реальному многообразию связей и процессов в меняющих численность популяциях. Именно их эколого-генетическая структура, опосредуя влияния разных факторов, определяет устойчивость популяций в сложнейших взаимодействиях факторов, внешних и внутренних по отношению к самой популяции.
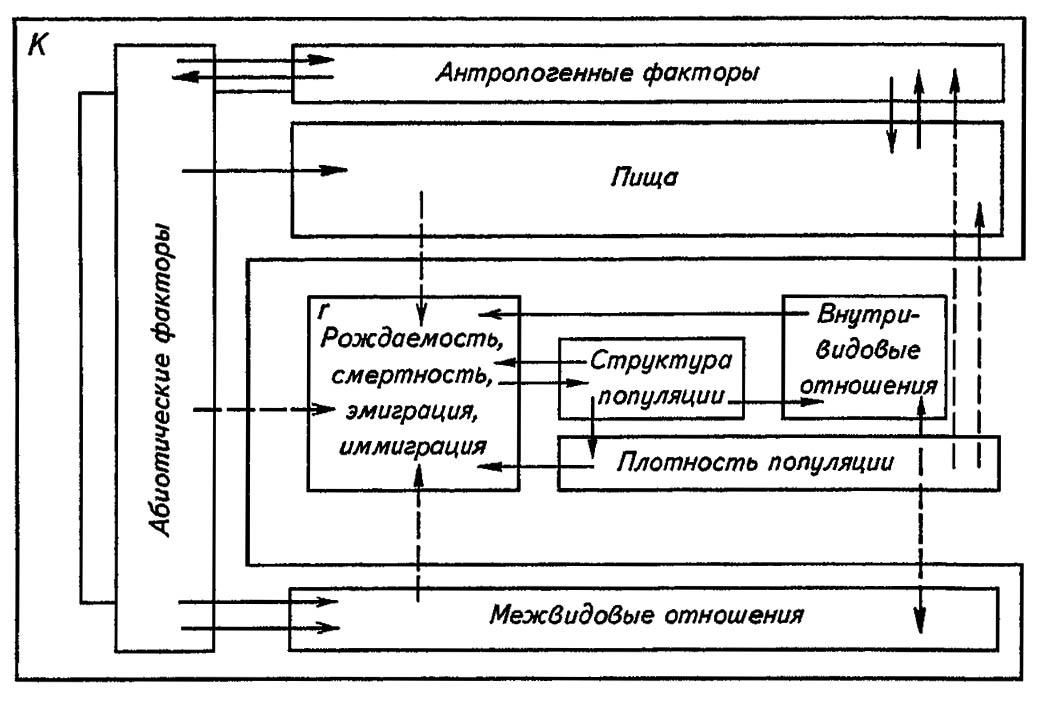
Рисунок 196. Факторы динамики численности популяций насекомых
Роль внутренних факторов ещё не оценена должным образом, однако очевидно, что понятие численности популяции неадекватно понятию её плотности. Эти понятия неразличимы в концепции автоматического регулирования. Плотность популяции приобретает особый смысл при подразделении факторов динамики численности на модифицирующие и регулирующие; при этом зависимость данных факторов от плотности популяции опосредуется её структурой. Таким образом, структура популяции становится фактором динамики численности.
Полагая, что численность насекомых всегда ограничивается некоторым сопротивлением среды и вместе с тем всегда сохраняется на уровне, достаточном для воспроизводства, мы противопоставили r-стратиотов, продуцирующих многочисленных потомков, не заботясь о достижении ими половой зрелости, и if-стратиотов, менее плодовитых, но проявляющих те или иные формы заботы о потомках, что позволяет им достигать половой зрелости. При освоении новых сред обитания, новых адаптивных зон преимущества имеют r-стратиоты, способные к быстрому наращиванию численности в отличие от менее плодовитых, но более конкурентоспособных r-стратиотов. Ведь в ещё не освоенных средах сопротивление среды незначительно. Однако в уже освоенных средах проявляются преимущества А-стратиотов: они постепенно вытесняют менее конкурентоспособных r-стратиотов.
С учётом приведённых рассуждений рассмотрим проблему вредоносности насекомых, так как практика сельскохозяйственного производства (распашка целинных земель, севообороты, сбор урожая, защитные мероприятия и тому подобное) обеспечивает постоянное обновление сред обитания, а проводимые защитные мероприятия постоянно нарушают естественный ход замещения видов и стабилизации энтомофауны в агроценозах. При этом создаются условия для роста численности r-стратиотов, что по установившейся традиции позволяет рассматривать вредоносность насекомых как прямое следствие их численности и плодовитости, обращаясь к таким элементам структуры популяций, как, например, распределение партий яиц, лишь во вторую очередь. Однако способность долгоносиков — цветоедов рода Anthonomus — распределять немногочисленные яйца по отдельным бутонам земляники (Anthonomus rubi Hbst.) или яблони (A pomorum L.) служит причиной высокой вредоносности этих видов при относительно малой численности и плодовитости.
Рассмотрим значение ещё одного аспекта обсуждаемой проблемы. Несмотря на исключительную прожорливость некоторых фитофагов и свойственную их популяциям способность к росту, существующие в природе виды растений не уничтожены ими полностью и в целом успешно противостоят нашествию вредителей. Анализ возможных факторов устойчивости растений на основе сопряжённой эволюции с насекомыми привлёк внимание исследователей к весьма разнообразным вторичным соединениям и метаболитам. Продуцируемые растениями эти соединения выполняют роль пищевых аттрактантов и стимулянтов или пищевых репеллентов и детеррентов для насекомых. О сложности взаимодействий с насекомыми можно судить, например, по тому, что выделяемый капустными (крестоцветными) растениями синигрин, служащий токсином и пищевым репеллентом для многих насекомых, приобретает значение жизненно необходимого пищевого стимулянта для капустной тли или капустной белянки: гусеницы последней предпочитают голодать и гибнут на всех необходимых для поддержания жизни субстратах, которые лишены синигрина.
Множество подобных примеров показывает значение для защиты растений от вредителей алкалоидов, флавоноидов, терпеноидов и других производимых растениями соединений, не связанных с их основным метаболизмом. Некоторые из этих соединений, образованные вследствие случайных мутаций и рекомбинаций, могут содействовать сокращению или утрате пищевой привлекательности растения, в котором они образовались. Такие растения, защищённые от растительноядных насекомых, входят в новую адаптивную зону, лишённую на некоторое время вредителей.
В ответ на формирующиеся физиологические барьеры растений насекомые-фитофаги могут, в свою очередь, подвергаться действию отбора, в результате которого образовавшиеся рекомбинанты и насекомые-мутанты проникают в новую для них адаптивную зону. Здесь в отсутствие конкурентов они свободно питаются и производят потомков.
В системе взаимодействий растение — насекомое роль более активного компонента, безусловно, принадлежит насекомому. Благодаря выраженной способности к развитию резистентности и различным механизмам детоксикации и депонирования токсинов, исключительной энергии размножения и скорости развития, способности легко приспосабливаться к новому корму популяции насекомых в эволюционной истории взаимодействий постоянно «преследуют» защищающиеся от них растения, и градации полифаг — олигофаг — монофаг, по-видимому, соответствуют уровни сопряжённости эволюционных преобразований.
Таким образом, сопряжённость эволюции с вырабатывающими средства противодействия растениями служит важным фактором динамики численности популяций насекомых, опосредуемым полиморфизмом, проявляющимся в выборе членами популяции, разных тенденций воспроизводства. Сопряжённость эволюции подтверждается также тем, что многие насекомые используют в качестве собственных гормонов и феромонов заимствуемые ими из растений стероиды.
Проведённая дифференциация факторов динамики численности и эволюционных тенденций воспроизводства популяций может стать определяющей при установлении общих подходов к защите сельскохозяйственных культур от вредителей, особенно при индустриальной технологии возделывания. Выделенные r-стратиоты ориентируют защитные мероприятия на регуляцию численности вредителей путём усиления неспецифичных по природе факторов смертности, Х-стратиоты — на регуляцию посредством специфичных факторов рождаемости (наводнение популяций стерилизованными самцами и мутантами, использование феромонов и гормонов, селекция устойчивых сортов с учётом биологической роли вторичных соединений и метаболитов).
Казалось бы, что в естественных биоценозах, лишённых сколько-нибудь явных антропогенных воздействий, динамика численности насекомых должна быть гораздо более стабильной, чем в агроценозах. Однако этому допущению противоречит массовое размножение фитофагов, которое, например, в сибирской тайге приобретает столь же грозный характер, как лесные пожары. Впрочем, многие исследователи полагают, что взрывы численности характерны лишь для немногих видов насекомых и имеют совершенно иную природу, нежели обычные популяционные волны, отмечаемые в агроценозах.
Наиболее сильны вспышки массового размножения сибирского шелкопряда (Dendrolimus superans sibiricus Tshtur.). Преодолевая мощность регулирующих факторов и нарушая тем самым действие стабилизирующих механизмов, вспышки массового размножения этого вида становятся кульминацией длительного процесса, который слагается из нескольких фаз (рис. 197).
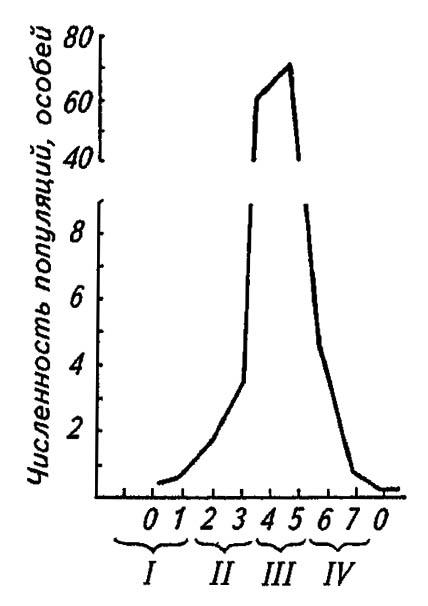
Рисунок 197. Последовательность фаз развития вспышки массового размножения насекомых (по Воронцову, 1975):
I — начало вспышки; II — продромальная фаза; III — эруптивная фаза; IV — фаза кризиса
В исходной фазе начала вспышки, или депрессии численности вредителя, она остаётся более или менее постоянной или при благоприятном сочетании экологических факторов возрастает в 2–3 раза, но не более.
В фазе подъёма, именуемой также продромальной фазой, численность и плотность популяций продолжают нарастать, а составляющие их особи отличаются от нормальных более интенсивным метаболизмом и повышенной двигательной активностью. Расселяясь на новые территории, они вступают в контакт и взаимодействие с другими, ранее изолированными группами. Все это приводит к резкому изменению структуры популяций, которые, выходя из-под контроля экологических факторов, обладают повышенными выживаемостью и генетическим потенциалом (рис. 198).
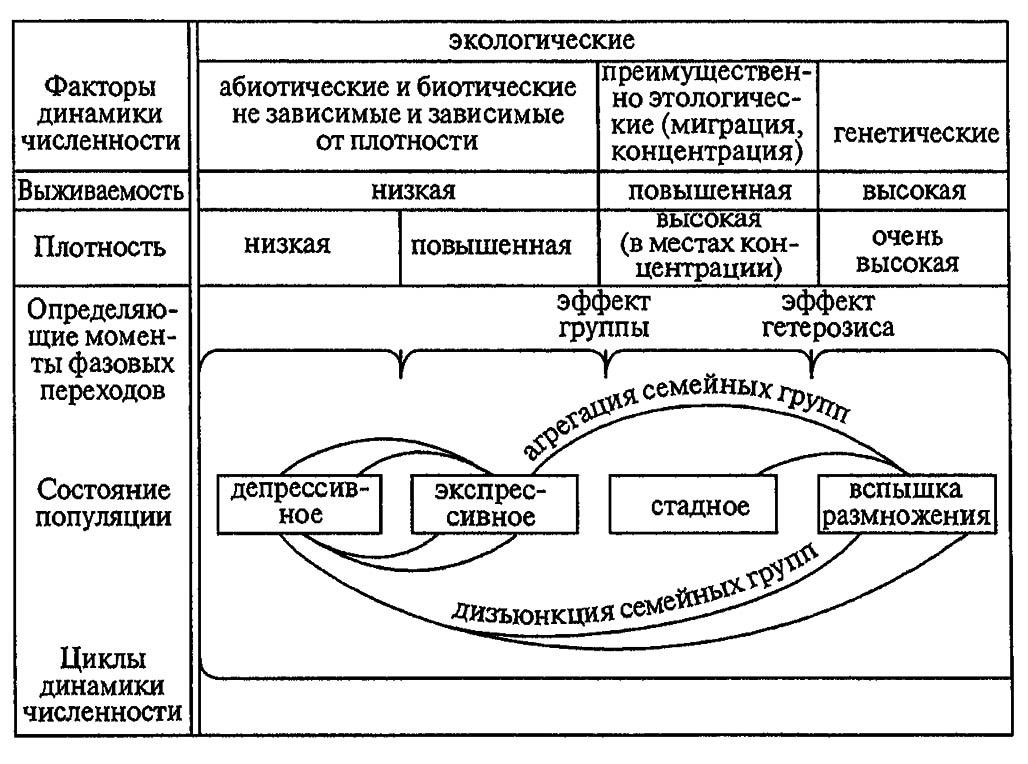
Рисунок 198. Схема взаимодействия факторов динамики численности популяций при вспышке массового размножения насекомых (по Рожкову, 1981)
В эруптивной, или фазе кульминации вспышки, причиняемый вред особенно значителен. Оголяя кроны деревьев, особи вредителя начинают испытывать дефицит корма, мигрируют на новые территории (в первую очередь самки) и при сохранении высокой плотности популяций в значительной степени подвергаются эпизоотиям.
Фаза спада, или кризиса, наступает не менее резко, чем взрыв численности. Происходит массовая гибель ослабленных и больных особей, что обусловливает резкое падение численности популяций и возвращение их под контроль регулирующих экологических факторов, вплоть до новой вспышки массового размножения насекомых.

