Климатические факторы
Среди факторов, составляющих климат Земли и отдельных её территорий, к основным по воздействию на живую природу можно отнести излучение Солнца, температуру и влажность атмосферы, к второстепенным — влияние ветра, ионизации и давления атмосферы, электрических полей.
Солнечное излучение
Все процессы в биосфере обеспечиваются излучением Солнца, но солнечная энергия распределяется по поверхности Земли неравномерно, и, кроме того, наблюдаются её сезонные и суточные колебания, особенно контрастные в умеренных и высоких широтах.
Определяя температуру воздуха и различных субстратов, солнечная радиация приводит к изменениям влажности и атмосферного давления. Проявления почти всех климатических факторов зависят от инсоляции и, в свою очередь, меняют интенсивность потока солнечных лучей над поверхностью почвы.
Действие света на насекомых проявляется, во-первых, через фотосинтез, в процессе которого создаётся органическое вещество, во-вторых, через изменения других экологических факторов. Однако примеров непосредственного воздействия света на жизнедеятельность насекомых сравнительно мало. Так, личинки стрекозы Aeschna umbrosa L. в темноте снижают темпы метаболизма и через некоторое время гибнут, а гусеницы Antheraea pernyi G.-М. реагируют на изменение светового режима изменениями в активности некоторых ферментов. Значение светового фактора для видов, активных только в ночное время, и обитателей пещер ограничено. Между тем яркий для глаза человека свет может быть тусклым для насекомых, но постоянное яркое освещение сказывается на их жизнеспособности, особенно на продолжительности жизни имаго. Более многочисленны примеры воздействия света на поведение и развитие насекомых.
Насекомые могут существовать более или менее длительное время и в полной темноте. Однако свет лежит в основе зрительных восприятий и различных биологических ритмов: суточных (циркадных), сезонных и лунных. Интенсивный ультрафиолетовый свет губителен для насекомых.
Температура
Температура — первичный периодический фактор, действующий на живой организм непосредственно и через изменения других факторов среды.
Свойства поверхности объекта — его цвет, структура, площадь — определяют интенсивность поглощения и излучения тепла, а масса объектов — их теплоёмкость. При уменьшении размеров объекта его теплоёмкость меняется быстрее, чем способность к восприятию и передаче тепла. Таким образом, при дефиците тепла в суровых условиях высоких широт более крупные организмы имеют некоторые преимущества по сравнению с мелкими.
В связи с тем что насекомые очень малы, они быстрее, чем другие животные, согреваются солнечными лучами, но быстрее и остывают в тени. Не имея постоянной температуры тела, они в значительно большей мере зависят от состояния среды, и излучение тепла для них существеннее, чем температура воздуха.
Диапазон температур, в котором возможны проявления активной жизнедеятельности насекомых, варьирует у разных видов. На графике, приведённом на рисунке 180, видно, что этот диапазон ограничен 15 и 38 °С, и за его пределами интенсивность жизненных процессов закономерно снижается.
Температурный оптимум, в зоне которого общие проявления жизнедеятельности наиболее эффективны при наименьших затратах энергии, смещён к повышенным температурам. Субоптимальные и супероптимальные температурные зоны допускают нормальную активность насекомых, но при дальнейшем изменении температуры наступает холодовое или тепловое оцепенение, ведущее к гибели. При тепловом оцепенении, ставшем постоянным, возврат к активной жизнедеятельности уже невозможен, но окоченение от холода сменяется переохлаждением до критической точки, когда освобождается скрытая теплота плавления и тело насекомого разогревается почти до 0 °С. Некоторые насекомые способны переносить весьма длительные и глубокие понижения температуры в состоянии анабиоза. Например, гусеницы кукурузного мотылька (Ostrinia nubilalis L.) оживали после охлаждения до —190 °С. При объяснении этого явления и при расчёте критической точки переохлаждения следует учитывать, что соки тела насекомого служат антифризами и замерзают при температурах более низких, чем вода.
В прикладных исследованиях часто приходится сталкиваться с необходимостью установления пределов выносливости и температур, предпочитаемых насекомыми. Взрослые особи непарного шелкопряда (Ocneria dispar L.), например, погибают при получасовом охлаждении до —4 °С. Имаго саранчи Melanoplus mexicanus L. гибнут при —8 °С через двое суток, а зимующие гусеницы златогузки: Nygmia phaeorrhoea L. способны выдерживать охлаждение до —14 °С в течение 158 дней.
При повышении температуры до 45–48 °С многие насекомые гибнут, но личинки некоторых жуков-златок и нимфы пустынной саранчи сохраняют активность при 52 °С, а личинки мух-береговушек активны даже при 65 °С.
Температуры, обычные для многих видов насекомых, весьма далеки от этих крайних значений. Способность насекомых к перенесению повышенных или пониженных температур варьирует в зависимости от физиологического состояния, предшествующего опыта и условий существования индивидов.
При выявлении предпочитаемых температур (термопреферендума) обычно используют приборы, поддерживающие градиент тепла или дающие возможность выбора одной из двух экспериментальных камер, в разной степени нагретых. В этих условиях комнатная муха (Musca domestica L.) обычно предпочитает температуру 42 °С, вошь Pediculus humanis L. — от 24 до 32 °С, но значения термопреферендума могут меняться в зависимости от сопутствующих обстоятельств. Например, муравьи Formica rufa L., содержавшиеся ранее при 3–5 °С, избирали температуру от 23 до 29 °С, а содержавшиеся при 27–29 °C — температуру от 32 до 52 °С. Для пустынной саранчи (Schistocerca gregaria Forsk.) выявлен следующий ход изменений термопреферендума, °С: нимфа I возраста — 30,1, II возраста — 28,8, III возраста — 31,6, IV возраста — 37,1, нимфа V возраста — 36,7, молодые имаго — 39,3, половозрелые особи — 29,4. В ночное время многие насекомые избирают более низкие температуры, чем днем.
В зависимости от термопреферендума, выражающего биологические требования вида, и от диапазона переносимых температур насекомые разделяются на стенотермных и эвритермных. Первые выносят лишь ограниченные колебания теплового режима, вторые способны переносить изменения фактора в более широких пределах. Паразиты теплокровных животных, как и многие представители фауны тропиков, как правило, стенотермные и теплолюбивые. Среди стенотермных холодолюбивых видов фауны снегов Гималаев выделяются группы ногохвосток и двукрылых, активных в диапазоне температур от 0 до 10 °С, от 0 до 5 °С и наконец активных только в солнечные часы дня при температурах от 5 до 10 °С. Представители двух первых групп настолько чувствительны к повышению температуры, что гибнут через несколько минут от тепла руки человека.
Примеры эвритермных видов: зелёная падальная муха (Lucilia serricata Mg.), бабочка-крапивница (Aglais urticae L.), панцирные клещи (Oribatei), обитающие на голых скалах внутриконтинентальных районов и переносящие колебания от 60 днем до 0 °С ночью.
Насекомым присуща и собственная терморегуляция. Об этом свидетельствует, например, то, что мёртвые жучки-чернотелки в пустыне нагреваются на 2–9 °С сильнее, чем живые. У чёрного таракана (Blatta orientalis L.) благодаря испарению с поверхности тела или усиленному потреблению кислорода температура либо слегка повышается, либо понижается, но только, если температура среды выходит за пределы 13–25 °С.
Медоносные пчелы в жаркую погоду проветривают улей, но при температуре ниже 13 °С начинают усиленно двигаться и скучиваются. При этом особи, остывающие на периферии скопления, переползают в его центр, вытесняя разогревшихся. В результате температура воздуха в улье повышается до 30 °С. Большое значение для терморегуляции имеет вибрация крыльев шмелей и бражников, которые способны взлетать лишь при температуре крыловых мышц не ниже 34,5 °С. Многие виды насекомых повышают свою температуру, выползая на освещённые солнцем места.
Для оценки влияний температуры на ход развития насекомых обычно пользуются критерием суммы эффективных температур, отсчёт которых ведут от заранее вычисленных значений нижнего порога развития. Например, муха-дрозофила (Drosophila melanogaster Mg.) развивается при температуре не ниже 13,5 °С — пороговой для данного вида. Соответствующие значения температурного порога для яблонной тли (Aphispomi Deg.), комнатной мухи (Musca domestica L.) и рыжего таракана (Blatella germanica L.) составляют 7,12 и 6,5 °C. Для амбарного долгоносика (Sitophilus granaria L.) температурный порог развития 11 °С, для капустной моли (Plutella maculipennus Curt.) — 9,8, шведской мухи (Oscinella frit L.) — 8, озимой совки (Agrotis segetum Schiff.) — 10, колорадского жука (Leptinotarsa decemlineata Say.) — 12°C.
При более низких температурах развитие тормозится, но если их среднесуточные значения превосходят пороговые, то развитие продолжается и насекомые через положенный срок достигают половой зрелости.
В допустимых для каждого вида пределах длительность развития сокращается с повышением температуры, а его скорость возрастает. Эта зависимость, выраженная гиперболой Блунка, соответствует принципу кинетики химических реакций, по которому увеличение температуры на 10 °С приводит к ускорению реакций и процессов в 2–4 раза. Так, эмбриональное развитие колорадского жука при 17 °С завершается через 216 ч, а при 27 °С — через 96 ч.
Способность биологических. объектов к терморегуляции, активному выбору предпочитаемых температур и поддержанию гомеостаза позволяет противопоставить термостабильные и термолабильные процессы, стадии и фазы, которые в меньшей или большей степени подвержены влияниям температуры. В частности, период дробления в эмбриогенезе колорадского жука относительно термостабилен, а его заключительные этапы термолабильны.
Способность широко распространённого в Европе клопа-солдатика (Pyrrhocoris apterus L.) давать только одно поколение и на юге, и на севере ареала определяется избирательностью субстратов, нагреваемых не более чем до 23 °С.
Расчёт сумм эффективных температур (S) проводят по формуле:
S = (t-t0)n,
где t — среднесуточная температура воздуха; t0 — значение нижнего порога развития; n — число дней, необходимых для развития.
Например, Drosophila melanogaster Mg. (t0 = 13,5 °C) при 26 °C развивается 20 дней, а при 19,5 °C — 41,7 дня. Таким образом, и в первом случае, когда S= (26 — 13,5)20 = 250, и во втором — S = (19,5- 13,5)*41,7 = 250,2 сумма эффективных температур одинакова (тепловая константа) и может быть использована для прогноза сроков развития, так как n = S/(t — t0).
Для яблонной тли сумма эффективных температур (S),°C — 114, комнатной мухи — 230, амбарного долгоносика — 360, капустной моли — 380, шведской мухи — 400, озимой совки — 1 000, для рыжего таракана — 1900. Зная, например, что в Ленинградской области сумма эффективных температур сверх порога развития капустной моли 9 °С составляет примерно 740 °С, можно ожидать появления здесь не менее двух поколений вредителя. В Омской области (S = 1 150 °С) возможно развитие трёх поколений этого вредителя, в Краснодарском крае (S = 2 300 °С) — уже 5–6 поколений. Такие расчёты применительны лишь к тем видам, число поколений которых варьирует в зависимости от условий существования и не закреплено генетически, как, например, у тутового шелкопряда Bombyx mori L.
Нередко регистрация сумм эффективных температур позволяет прогнозировать сроки появления вредящих фаз и планировать защитные мероприятия. В частности, инсектициды против колорадского жука на картофеле целесообразно применять при S = 150 °С (при t0 = 12 °С), поскольку к этому времени начинается массовое появление вредителя.
Расчёты сумм эффективных температур имеют ориентировочное значение; их следует корректировать с учётом данных непосредственных наблюдений за состоянием популяций вредителей. Кроме того, сроки развития вредителей зависят не только от накопленного тепла, но и от колебаний температуры, которые обычно содействуют их сокращению. При пониженных температурах длительность жизни отдельных особей значительно возрастает. Так, если при 30 °С мухи Drosophila melanogaster Mg. существуют в среднем 21 день, то при 15 °С — не менее 130 дней.
Подвержена влиянию климатических факторов и плодовитость насекомых: она максимальна при оптимальных условиях, но пожизненная плодовитость самок зависит от сроков их существования. Нередко сокращение числа яиц, откладываемых самкой ежедневно при пониженных температурах, компенсируется длительностью периода их откладки, и в конечном счёте максимально плодовитыми оказываются наиболее долговечные, а не наиболее активные особи. Более всего яиц в единицу времени самки шведской мухи откладывают при температуре 18–39 °С, муха-меромиза (Meготуza saltatrix L.) — при 20–30, а луговой мотылёк Loxostege sticticalis L. — при 25 °С.
Влажность
Отделить влияние влажности от воздействия других экологических факторов трудно, а порой и невозможно. Если имеются доступные источники влаги, насекомые легко переносят сухость воздуха, и лишь формы, постоянно обитающие в водоёмах (гидробионты), гибнут на суше.
Влияние влажности на длительность и скорость развития обычно определяется воздействием температуры. У комнатной и зелёной падальной мухи (Musca domestica L. и Lucilia serricata Mg.) обнаружена линейная зависимость между скоростью развития и дефицитом влажности, а у перелётной саранчи скорость развития возрастает при повышении относительной влажности до 10 %. При более высокой влажности развитие саранчи тормозится и многие особи поражаются грибными и бактериальными болезнями.
Сухопутных насекомых подразделяют на гигрофилов, обитающих во влажных местах, мезофилов, менее требовательных к влажности атмосферы, и ксерофилов, приспособленных к постоянному дефициту влажности.
Для пополнения запасов воды некоторые насекомые пьют росу (клоп-черепашка — Eurygaster integriceps Pot.) или соки растений, потребляют влагу с пищей либо используют метаболическую воду, образуемую при окислении жиров и других органических веществ. Гусеницы платяной моли (Tineola biselliella L.) и мельничной огнёвки (Ephestia kuhniella Zell.), амбарный и рисовый долгоносики (Sitophilus granaria L. и S. oryzae L.), питаясь субстратами с небольшим содержанием влаги, обходятся метаболической водой. Между тем гигрофильные насекомые, и в особенности дышащие жабрами гидробионты, потребляют значительное количество воды через проницаемые покровы.
Один из способов экономии водных запасов, постоянно расходуемых при обмене веществ, транспирации и испарении, — выделение концентрированной мочи. Для выведения избытка воды, потреблённой с соком растений, служат фильтрационные камеры кишечника.
Снижение содержания воды в организме разные виды насекомых переносят неодинаково: колорадский жук (Leptinotarsa decemlineata Say.) выдерживает подсушивание на 50 %, жук Catalpa lanigera L. — на 25%, а жук Lachnosterna sp. погибает при подсушивании на 15 %. Между тем личинки некоторых комаров, живущие в пересыхающих лужах, остаются живыми в течение 10 лет, сохраняя не более 3 % воды от массы тела. Однако эти данные характеризуют разнообразие реакций насекомых на изменение влажности лишь в экспериментальных условиях.
В природе влияние влажности опосредуется температурой и другими факторами среды. Поэтому в практике экологических исследований обычно рассматривают совместное влияние влажности и температуры, для демонстрации которого используют многообразные индексы, коэффициенты и диаграммы. При их расчёте и построении удобно пользоваться принятыми в метеорологии параметрами: усреднёнными (среднедневными, среднедекадными и тому подобными) значениями температуры воздуха и влажности, причём влажность учитывают либо в относительном выражении, либо в виде суммы осадков за тот или иной период.
Для прогноза и выявления лимитирующих факторов применительно к каждой стадии развития насекомого используют биоклимограммы Б. П. Уварова (рис. 181).
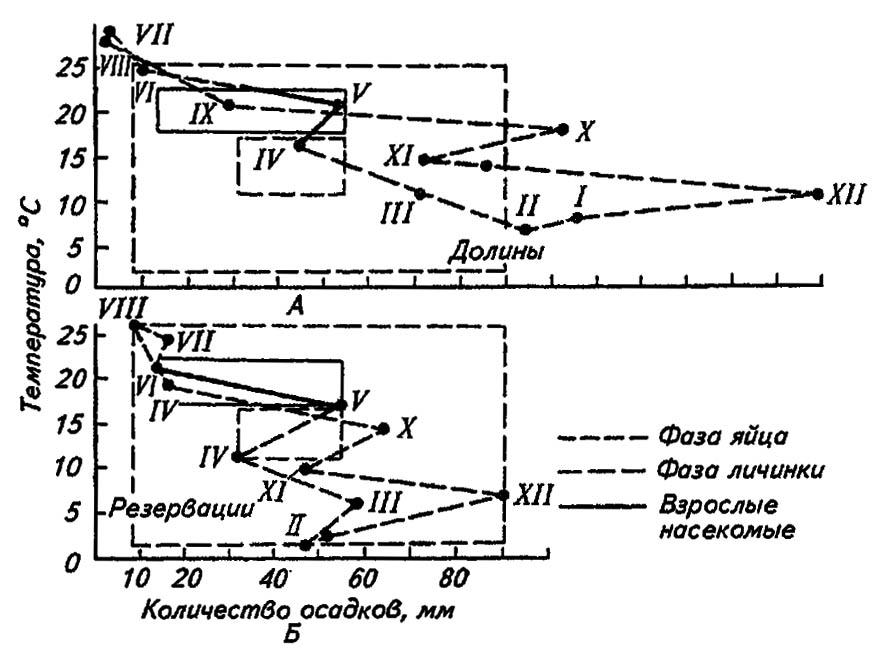
Рисунок 181. Биоклимограмма Б. П. Уварова для мароккской саранчи в Малой Азии (по Бей-Биенко, 1966):
А — неблагоприятные и Б — благоприятные условия жизни
Располагая метеосводками для территории и сезона массового появления вида, на соответствующем графике можно очертить зону оптимума, контуры которой при наложении на графики, построенные для иных территорий и сезонов, сразу выявляют, что и для какой фазы развития служит лимитирующим фактором. Из приведённого графика видно, что избыточные осадки в октябре (X), декабре (XII), январе (I) и феврале (II) были причиной гибели зимующих яиц мароккской саранчи, как и их недостаток и повышение температуры в летнее время (VI–VIII). Понижение температуры в апреле (IV) неблагоприятно сказывалось на имаго.
Второстепенные факторы
При сильном ветре насекомые, не успевшие укрыться в убежищах, разносятся на далёкие расстояния. Оказавшись в непривычных для себя условиях, они гибнут или приспосабливаются к этим условиям, причём эвритопные виды, отличающиеся повышенной экологической валентностью, имеют больше шансов на выживание, чем стенотопные виды. Нечасто условия на новой родине благоприятны для мигрантов, и насекомые обычно избегают порывов ветра. Все же некоторые из них, обладая своеобразными парусами — пушистыми щетинками и выростами тела, приспособлены к пассивному расселению токами воздуха.
При резком падении атмосферного давления, что обычно предвещает бурю, насекомые особенно активны. В этом нетрудно убедиться, наблюдая за муравейником. Позднее, прекращая перемещения и снижая активность, они пережидают ураган, но остаются подверженными косвенным влияниям ветра, меняющего температуру и содействующего испарению влаги. Пониженное атмосферное давление стимулирует окрыление некоторых видов бабочек и веснянок. Кроме того, насекомые при миграциях как бы следуют за снижением давления, устремляясь в те районы, где значения его ниже. Они также восприимчивы к ионизации атмосферы до и после грозы, во время лесных пожаров и электрических разрядов молний.
Положительные ионы, как правило, стимулируют активность насекомых, к отрицательным они безразличны, но в лабораторных экспериментах тли Acyrthosiphon pisi Kalt. реагировали на них торможением линьки. На многих насекомых влияет изменение направленности электрического поля, вероятно, в связи с накоплением электростатических зарядов на теле. Во время геомагнитных бурь активность многих насекомых повышается, а дрозофилы реагируют даже на слабое электрическое поле (до 200 В/м) и его изменения, например, при прохождении над ними облаков.

