Микроклимат и регуляция жизненных циклов
Совместное влияние климатических факторов определяет условия существования и жизнедеятельности насекомых в самых общих чертах. В этом смысле можно утверждать, что климат Заполярья, например, слишком суров для колорадского жука, вредоносность которого проявляется в более тёплых районах.
Наряду с метеорологическими условиями для насекомых ввиду их малых размеров и подвижности особое значение приобретают микроклимат, то есть климат на уровне организма, реальные условия существования, которые определяют биологические реакции насекомого в данное время и в данном месте.
Обычные агротехнические приёмы существенно меняют микроклимат пашни, который становится более суровым после жатвы и вспашки. При этом влажность воздуха в поле редко падает ниже 50 %, а для обитающих на растениях насекомых она более высока и постоянна. В целом микроклимат пашни характеризуется большими колебаниями температуры, повышенной влажностью, ослабленными порывами ветра и осадками, бедностью красными и синими лучами солнечного спектра, но относительным богатством инфракрасными.
Пределы изменчивости климатических (микроклиматических) факторов, которые насекомые переносят в недеятельном состоянии, значительно шире пределов их активности, а тем более пределов развития и размножения. Нередко виды, весьма устойчивые к различным климатическим воздействиям среды, после холодной зимы и жаркого лета устремляются в брачный полет только ранней осенью, в предвечерние часы редких тёплых и ясных дней.
Таким образом, при воспроизводстве популяций диапазон допустимой изменчивости факторов ещё более ограничен.
Зависимость размножения и развития насекомых от внешних условий корректируется и опосредуется соответствующими актами поведения. Обычно любое изменение воздействия фактора вызывает некоторое повышение активности и ориентировочную реакцию, направленную на отыскание благоприятных условий. Однако при невозможности их выбора насекомые впадают в оцепенение, когда резко сокращается обмен веществ, или вырабатывают ещё более совершенные и специфические приспособления.
В частности, некоторые многохвостки испытывают нимфоз — изменение облика, сопутствующее сокращению активности. Втягивая придатки и облекаясь дополнительной линочной шкуркой, они становятся похожими на мелких куколок высших насекомых и в этом состоянии пребывают до наступления благоприятных условий существования. Для личинок большого мучного хрущака (Tenebno molitor L.) обычно явление реверсии, связанное с ухудшением условий существования: продолжая линять, они сокращаются в размерах и начинают расти лишь в благоприятных условиях.
Однако наиболее распространённое средство перенесения экстремальных условий среды — диапауза — долговременное торможение метаболизма и всех видов активности в ответ на сигнальные влияния сезонных изменений климата. Диапауза предваряет наступление зимней бескормицы и холодов или иссушающего летнего зноя. Обычный стимул к ее развитию — астрономически точное изменение фотопериода.
В момент наступления диапаузы и температура, и количество корма вполне отвечают нормам поддержания активной жизнедеятельности насекомого, но диапауза наступает в ответ на сокращение продолжительности дня — признак приближения осени. Если нарушить естественный режим освещённости в конце лета, осветив плодовый сад на 2 мин после захода солнца, то вредоносные гусеницы листовёртки Adoxophyies sp. не впадают в диапаузу и погибают при первых же заморозках. Таким образом, диапауза не сводится к оцепенению и другим формам физиологического покоя, наступающим при воздействии экстремальных условий. Более того, для её преодоления необходимо длительное воздействие пониженных температур. В противном случае, например, если до завершения диапаузы перенести насекомое в тёплые условия, оно вскоре погибнет.
Для многих обитателей умеренных широт с резко выраженной цикличностью сезонных изменений климата диапауза становится необходимым элементом жизненного цикла, обеспечивающим его продолжение после зимовки. Содействуя развитию особой устойчивости к пониженным температурам, она вместе с тем обеспечивает экономное расходование накопленных ранее резервов и синхронизацию потребностей насекомого с возможностями их удовлетворения. Для обитателей тропиков диапауза — средство защиты от перегревания и потери влаги в период сухого сезона.
Способность впадать в диапаузу закреплена генетически, но её реализация и сроки программируются фотопериодом. Например, у тутового шелкопряда она наступает в фазе яйца, но только в том случае, если отложившая его самка развивалась при строго определённом режиме освещённости. При этом в её подглоточном ганглии синтезируется специфический гормон, проникающий в формирующиеся яичники и достигающий ооцитов. У повреждающей хлопчатник хлопковой моли (Pectinophora gossypiella St.) гусеницы последнего возраста впадают в предкуколочную диапаузу в зависимости от фотопериода в момент инкубации яиц, а у колорадского жука имагинальная диапауза наступает вскоре после окрыления. Во всех этих случаях для фотопериодической реакции имеет значение лишь абсолютная длительность светлого времени суток, но у стрекозы Anax imperator L. диапауза возникает лишь при удлинении дня со скоростью менее 2 мин в сутки. Достигая последнего возраста в начале июня, когда день нарастает быстрее, стрекоза окрыляется в тот же год, однако те особи, которые задерживаются в развитии, окрыляются весной следующего года, после диапаузы.
Подавляющее большинство насекомых активно при длинном дне и уходят в диапаузу при сокращении фотопериода (рис. 182, А). При этом даже сумеречная освещённость воспринимается как продолжение дня и предотвращает диапаузу. У некоторых обитателей тропиков, наоборот, фотопериодическая реакция проявляется при удлинённом дне, и, таким образом, по активности они короткодневные (рис. 182, Б). Среди них наиболее известен тутовый шелкопряд. Кукурузный мотылёк, капустная и репная белянки и некоторые другие виды диапаузируют при средних значениях фотопериода, оставаясь активными при длинном и коротком дне.
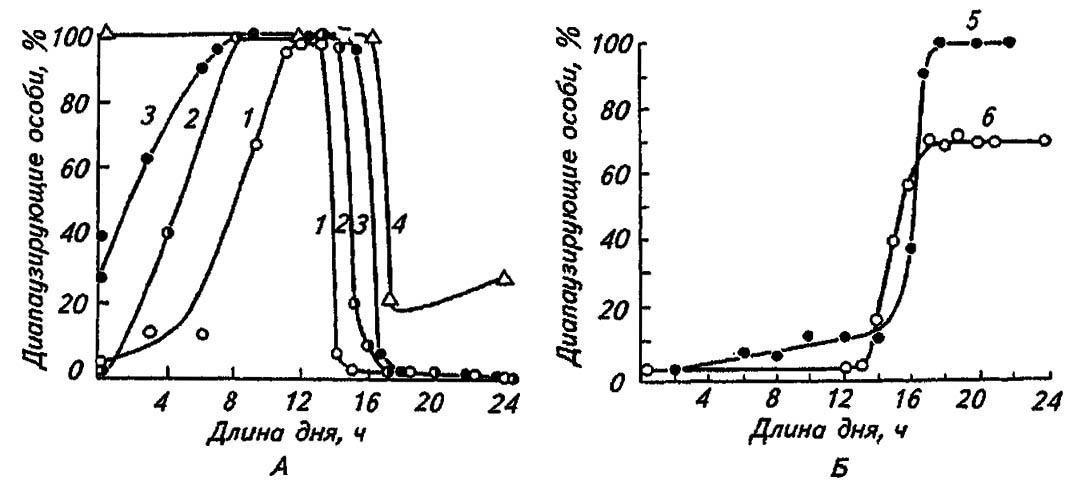
Рисунок 182. Изменение числа диапаузирующих особей под влиянием фотопериода у длиннодвевных (А) и короткодневных видов (Б) (по Данилевскому, 1965):
1 — восточная плодожорка; 2 — капустная белянка; 3 — совка Acronycta rumicis; 4 — колорадский жук; 5 — цикадка Stenocranus minutus; 6 — тутовый шелкопряд
Приведённые графики демонстрируют, кроме того, гетерогенность популяций и пределы индивидуальной изменчивости в фотопериодических реакциях. Например, у восточной плодожорки (Grapholitha molesta Busk.), относимой к длиннодневным видам, некоторая часть популяции впадает в диапаузу при сокращённом фотопериоде.
С ещё большей определённостью лабильность фотопериодических реакций проявляется у видов, распространённых повсеместно, например у совки Acronycta rumids L. Будучи длиннодневной формой на широте Санкт-Петербурга, она уходит в диапаузу при 19-часовом освещении, а на побережье Чёрного моря — при 15-часовом. При повышении температуры наблюдается смещение порогов фотопериодической реакции к некоторой короткодневности (рис. 183), что соответствует зональным изменениям климата.
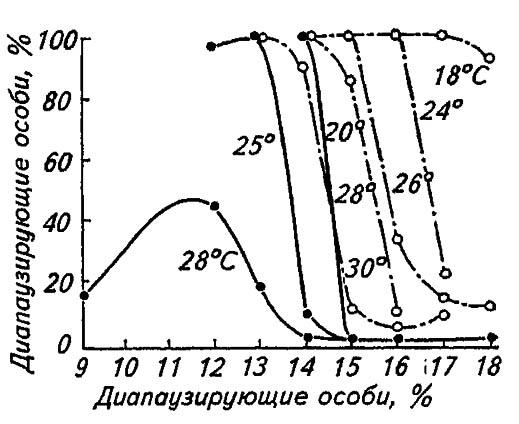
Рисунок 183. Сдвиг фотопериодической реакции под воздействием различных температур (по Данилевскому, 1965):
сплошные линии — хлопковая совка; прерывистые — колорадский жук
В развитии диапаузы выделяют подготовительный период, когда испытавшие фотопериодическую индукцию насекомые накапливают необходимые резервы, тормозят половую активность и резорбируют сформированные ранее яйца. Далее происходит общее подавление активности и газообмена, обезвоживание тканей тела и дегенерация крыловых мышц. Все эти процессы контролируются нейросекретами, переводящими метаболизм на гликолитический путь, и через 2–3 недели подготовительный период сменяется периодом диапаузы. Покоящиеся в этом состоянии насекомые отличаются высокой устойчивостью к экстремальным условиям существования.
Через 1,5–2 мес или более начинаются процессы реактивации, которые при благоприятных условиях среды приводят к полному восстановлению активности. Однако у многих насекомых реактивация тормозится низкими температурами в конце зимы, и состояние диапаузы постепенно переходит в состояние физиологического покоя — оцепенение, которое может быть прервано в любое время.
Изредка сильные оттепели в конце зимы провоцируют выход насекомых с мест зимовки, что может повлечь за собой их гибель от переохлаждения. Многие аспекты и процессы диапаузы насекомых остаются ещё неясными, поэтому приведённые описание и схема (рис. 184) в дальнейшем должны быть конкретизированы.
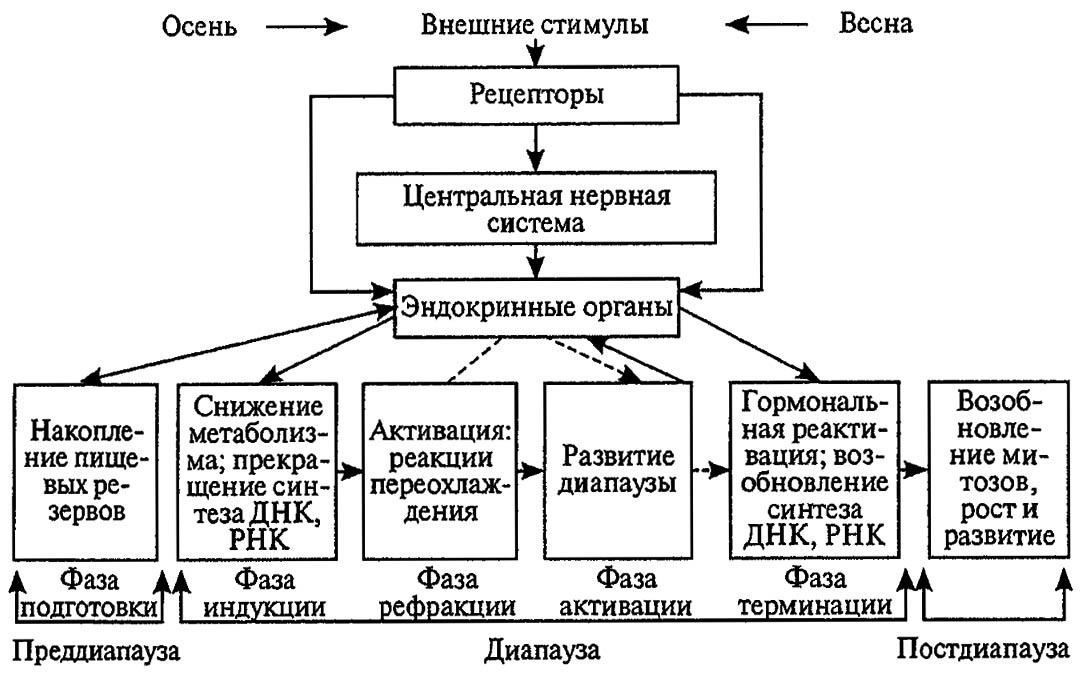
Рисунок 184. Последовательные фазы развития диапаузы у насекомых (по Gillot, 1980)
О сложности диапаузы можно судить по состояниям колорадского жука, способного впадать в зимнюю и летнюю диапаузу, испытывать повторную и затяжную диапаузу (суперпаузу), длящуюся несколько лет, или временно прерывать свою активность, впадая в разные формы физиологического покоя (рис. 185).
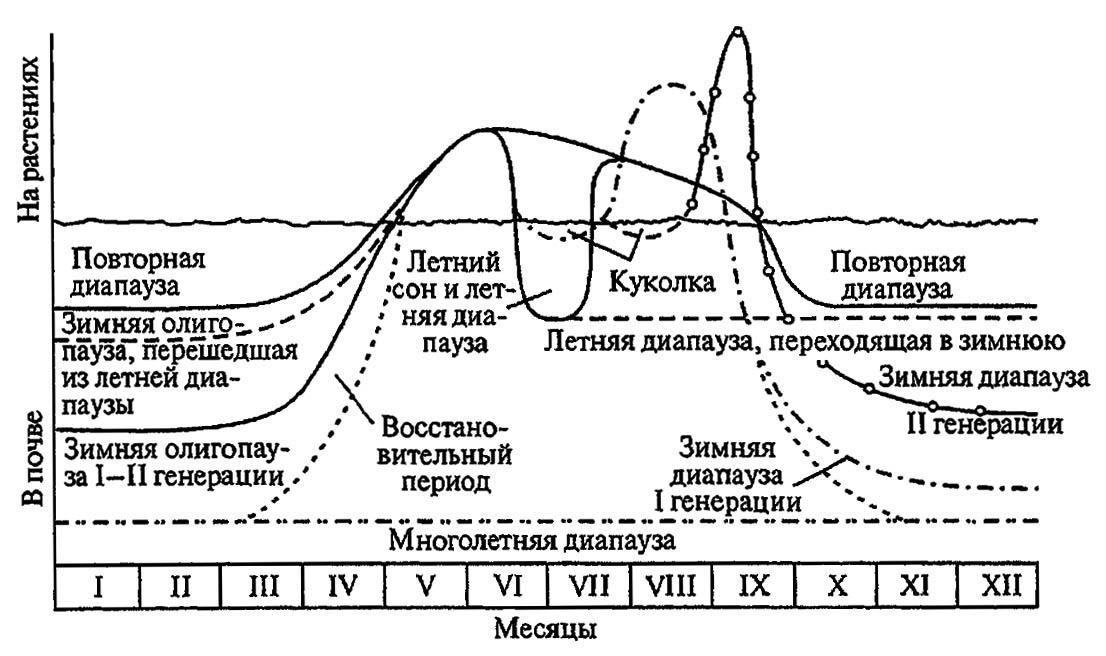
Рисунок 185. Различные типы диапаузы у колорадского жука (по Ушатинской, 1982)

