Свойства популяции насекомых
Популяция — это естественное множество особей, объединённых генетическими и экологическими связями. Констатация генетических связей предполагает принадлежность особей к одному виду и способность скрещиваться друг с другом, а экологических связей — сосуществование видов в определённых стациях при определённом отношении к среде. Популяция — не случайное скопление особей одного вида, а естественное множество со своей структурой и целостностью, проявляющейся во взаимодействиях с другими популяциями и видами. Подчиняясь эндогенным стимулам и сигналам внешнего мира, популяции способны распадаться на дочерние, сливаться с другими популяциями, восстанавливать и утрачивать свою целостность, но всегда в закономерной последовательности состояний и фаз, в регулярных изменениях численности, ограниченных лишь возможностью воспроизводства и расселения.
Популяции можно характеризовать по генофонду, возрастной структуре, плотности распределения особей и размещению их в стациях, соотношению рождаемости и смертности, динамике численности и способности к воспроизводству.
Размещение насекомых
Насекомые размещаются по различным средам в соответствии с требованиями к условиям существования и с инстинктом продолжения рода. Строгая приуроченность к конкретным местообитаниям отмечена у видов с узкой экологической валентностью — стенотопов и монофагов. Эвритопные виды встречаются в большем наборе сред, а виды, именуемые убиквистами, вообще не имеют приверженности к каким-либо местообитаниям. Например, семиточечную божью коровку (Coccinella septempunctata L.) можно найти и на пшеничном поле, и под пологом леса, на болотах, и в больших городах на асфальте, и на стенах зданий. Напротив, некоторые пухоеды и вши паразитируют только на «своих» видах птиц и млекопитающих, избирая при этом лишь отдельные части их тела.
При описании размещения отдельных особей обычно пользуются термином «местообитание», но для популяций и вида в целом употребляют термины «стация» и «ареал». Ареал включает в себя стации всех популяций данного вида, а стация — все местообитания особей, образующих популяцию.
В отношении занятых территорий уместно противопоставление аборигенных видов—автохтонов, существующих в данном месте изначально, и аллохтонов — пришельцев, прибывших из других мест. Эти противопоставления существенны при сравнении разных фаун, например фауны Западной Европы и Северной Америки. Около половины (45 %) обычных для этих фаун видов — аллохтоны, причём в Америку из Европы было завезено в 10 раз большее их число. Многие насекомые вскоре проявили себя как опасные вредители сельскохозяйственных культур, так как на новом месте они избавились от прежних хищников и паразитов — автохтонов Европы. Однако и завезённый в Европу колорадский жук до сих пор не имеет эффективных местных паразитов и хищников.
Различают виды-эндемики, которые существуют только в данном месте и нигде более, и виды-космополиты, распространённые повсеместно. Их ареал не ограничен.
Приверженность многих видов насекомых избранным стациям и местообитаниям отражена в их характеристиках и названиях. Например вредящие посевам Agrotis segetum Schiff. и Anisopha segetum Hbst. именуются соответственно посевной совкой и посевным хрущом; тополевый листоед Melasoma populi L. обитает на тополе, а близкий вид Melasoma tremulae F — на осине (Populus — тополь, Populus tremula — осина) Известная всем комнатная муха Musca domestica L. (domestica — домовая) обитает в жилищах человека, а желудочный овод (Gastrophilus intestinalis DeG.) (gaster — желудок, intestinum — кишечник) паразитирует в пищеварительном тракте млекопитающих.
Свойства видов избирать определённые стации отражены в сформулированном Г. Я. Бей-Биенко принципе стациальной верности и логически связанном с ним принципе зональной смены стации. Действие этих принципов можно проследить на следующих примерах. Если популяции перелётной саранчи на севере ареала приурочены к песчаным, хорошо прогреваемым солнцем почвам, то на юге они стремятся в сырые, болотистые места. При этом их потребность в определённых гидротермических условиях в связи с зональными изменениями климата и почв приводит к смене стации. Восточный майский хрущ (Melolontha hippocostani F.) заселяет опушки леса и поляны на севере ареала, но уходит под полог леса на юге; посевная совка Agrotis segetum Schiff. сменяет пары и посевы озимых культур на орошаемые поля. Выраженные в этих примерах принципы отражены на схеме, показанной на рисунке 189: переходу от лесной зоны к зоне полупустынь соответствуют изменения стаций — от относительно ксерофитных к мезофитным и гигрофитным.
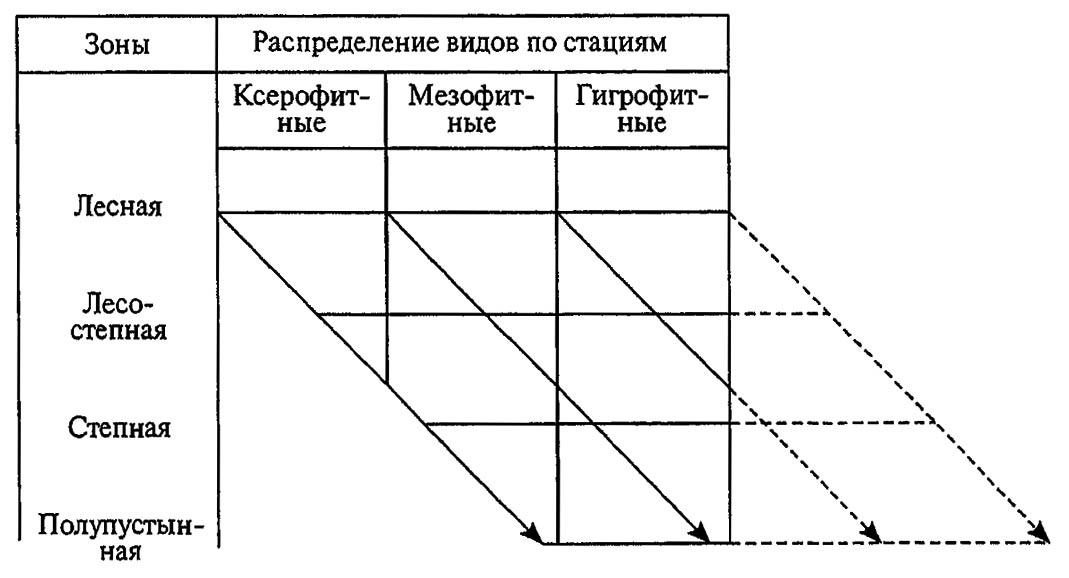
Рисунок 189. Диаграмма зональной смены стаций (по Бей-Биенко, 1966)
Сходные изменения можно отметить на фоне сезонной периодичности уровней влажности и температуры, которой соответствует сезонная смена стаций. Зональная смена ярусов отмечена, например, у короеда Blastophagus piniperda L. Обитая под корой сосны на севере, этот вредитель переходит на её корни на юге. При движении с севера на юг можно наблюдать, что личинки жука-оленя (Lucanus cervus L.) меняют стволы деревьев на корни, а пшеничный трипс Haplothrips tritici Kurd, от зимовки в стерне злаков переходит к зимовке в почве, сохраняя приверженность не стации, а определённым режимам влажности и температуры, сравнимым в разных стациях различных зон.
Таким образом, выбор насекомыми конкретных сред обитания не случаен и определяется многими обстоятельствами. Однако в пределах стаций, достаточно однородных на всем протяжении, отдельные члены популяции распределяются иногда без видимого порядка.

