Биотические факторы
Особенность биотических факторов среды состоит прежде всего в том, что они взаимодействуют с подверженными их влиянию популяциями и зависят от их свойств. Если одни из этих факторов жизненно необходимы для насекомых (симбионты, половой партнёр, источник пищи и другого), то действие других (конкуренты, паразиты, хищники и др.), напротив, отрицательно. Однако хищники, например, могут косвенным образом содействовать благополучию популяций за счёт уничтожения ослабленных и больных особей.
Многие хищники и паразиты реагируют на состояние популяций, ослабляя или усиливая своё действие двумя принципиально различными способами. При возрастающей плотности популяции жертв они, сохраняя свою численность неизменной, становятся более прожорливыми и агрессивными. Этой функциональной реакции можно противопоставить реакцию численную, когда плотность популяции жертв стимулирует размножение и рост численности хищников и паразитов.
Взаимодействия биотических факторов с популяциями насекомых могут быть более или менее тесными. Диапазон этих взаимодействий включает экологические связи между членами одной и той же популяции, между разными популяциями одного вида и между разными видами. Противопоставляя внутривидовые и межвидовые отношения, оценим их влияния и результаты по изменениям темпов воспроизводства популяций, а также баланса между рождаемостью и смертностью.
Внутривидовые отношения
Наиболее явный пример внутривидовых отношений — взаимодействия половых партнёров, которые были рассмотрены выше. Здесь остановимся на обсуждении эффекта группы, массового эффекта и отдельных примеров внутривидовой конкуренции.
Эффект группы
Объединение насекомых в группы нередко содействует их выживанию и размножению. Роение самцов комаров служит средством привлечения самок, а совместное обитание личинок мух на трупах и экскрементах благодаря выделяемым ими протеолитическим ферментам способствует разжижению и усвоению субстратов. Особенно наглядно проявляется эффект группы при фазовом полиморфизме, свойственном саранчовым, гусеницам некоторых бабочек, немногим жукам, тлям, сеноедам, тараканам и сверчкам. Объединение в стаи часто провоцирует усиление метаболизма и активности насекомых, содействует их миграциям и расселению на новые территории, может вызывать вспышки массового размножения вредителей.
Массовый эффект
В отличие от эффекта группы массовый эффект, вызванный перенаселением среды, часто обусловливает сокращение популяций. Массовый эффект можно наблюдать на примере вредителей запасов, заселяющих элеваторы и мукомольные предприятия. Малый мучной хрущак (Tribolium confusion Duv.), зерновой точильщик (Rhizopertha dominica F.) и рисовый долгоносик (Sitophilus oryzae L.) сокращают плодовитость после того, как плотность их популяций достигнет некоторого предела. Одновременно замедляется развитие личинок, а особи Т. confusum Duv. начинают пожирать собственных потомков. Так, при числе жуков в 1 г муки, составляющем 1,25; 2,5; 5; 10; 20 и 40 особей, процент съеденных яиц с 7,7 возрастает соответственно до 17; 20; 39,7; 70,2 и 98,4.
Явления такого рода, названные самоограничением популяций, препятствуют их слишком быстрому вымиранию в ограниченных средах. Проявления массового эффекта отмечают до наступления конкуренции за пищу и жизненное пространство.
У паразитических хальцид Nasonia vitropennis L., заселяющих мух Phaenicia sericata Zt, сокращается способность к воспроизводству, когда плотность популяции паразитов становится чрезмерной. Это связывают с тем, что при малой численности хозяев происходит торможение откладки яиц, а отложенные яйца, оставаясь неоплодотворёнными, развиваются в самцов. Кроме того, гибель хозяев, заражённых более чем одной личинкой паразита, также приводит к возрастанию смертности личинок.
Внутривидовая конкуренция
Конкурентные отношения между особями одного вида проявляются в территориальном поведении, содействующем равномерному использованию имеющихся ресурсов, во внутрипопуляционной иерархии, а также в некоторых особых формах взаимодействия особей.
Муравьи и термиты обычно избегают устраивать свои гнезда в непосредственной близости друг от друга и агрессивны к пришельцам из других семей. Ожесточённые схватки скарабеев (Scarabaeus sacer L.), отбивающих друг у друга тщательно выкатанные навозные шары, и стычки личинок австралийского навозника (Aphodius howitti Hbst.) содействуют рассредоточению популяций. Сходные функции имеют некоторые песни прямокрылых, охраняющих занятую территорию. Равномерное использование ресурсов проявляется и в стремлении паразитов избегать ранее заселённых хозяев.
Внутрипопуляционная иерархия, предполагающая выделение доминирующих и подчинённых особей, наблюдается среди личинок майского хруща (Melolontha melolontha L.). Развиваясь в течение трёх лет, личинки дифференцируются по возрастным группам, причём старшие оказывают угнетающее воздействие на младших. Поэтому вылет взрослых жуков имеет явственные трёхлетние циклы, тогда как другие насекомые, развивающиеся столь же долго, но не конкурирующие между собой, окрыляются ежегодно.
Особыми формами внутривидовой конкуренции обладают личинки паразитических перепончатокрылых, существующие совместно в одном хозяине. Здесь некоторые из них преобразуются в мускулистых, снабжённых крепкими челюстями особей, лишённых, однако, кишечника. Разрывая и перемешивая ткани хозяина, они облегчают питание нормальных личинок, но сами через некоторое время погибают. К явлениям той же природы можно отнести формирование трофических яиц некоторыми муравьями. Предназначенные для выкармливания личинок царской пары, они с самого начала остаются стерильными.
Отношения такого рода необходимы для успешного существования и воспроизводства популяций — жертвуя в малом, они приобретают стабильность численности и устойчивость к внешним воздействиям.
Межвидовые отношения
Влияния, оказываемые разными видами друг на друга, могут быть положительными и отрицательными, обоюдными и односторонними. Различают отрицательные формы взаимодействий между видами, к которым относятся аменсализм, конкуренция, хищничество и паразитизм, а также положительные формы — мутуализм, синойкия, комменсализм, сотрудничество.
Конкуренция
Конкурирующие виды противодействуют друг другу в борьбе за пищу, укрытия, места откладки яиц. Следует отличать непосредственные влияния (аменсализм), когда присутствие одного вида невыносимо для другого из-за выделяемых им метаболитов или из-за агрессивных форм его поведения, от конкуренции, или соперничества, за источники существования и воспроизводства.
При активном соперничестве, именуемом также интерференцией видов, один из них лишает другого доступа к источникам пищи и возможным местообитаниям. Нередкое для стрекоз стремление во что бы то ни стало изгнать особей другого вида с выбранных ими мест посадки у водоёмов — один из примеров такого рода. Другой пример — трогательная охрана муравьями тлей, выражающаяся в том, что они отгоняют от тлей паразитов и хищников.
Пассивная конкуренция, или эксплуатация, развивается при совместном использовании ресурсов. В этом случае более конкурентоспособный вид постепенно вытесняет соперника. В основе этих взаимодействий лежит принцип конкурентного исключения, сформулированный Г. Ф. Гаузе. Суть принципа в том, что два вида, имеющие сходные требования к условиям существования, совместно обитать не могут. Один из них вытесняет другого.
Периодически изменяя микроклимат, можно добиться, например, устойчивого равновесного сосуществования популяций хрущаков Tribolium castaneum L. и Т. confusum Duv., имеющих сходные потребности в пище, но различающихся по отношению к влажности и температуре. Первый вид явно предпочитает условия тропиков, второй — аридные условия.
Весьма показательны отношения трёх североамериканских видов рода Megarhyssa, откладывающих яйца в личинок пилильщика Tremex columba Panz. Сосуществуя и имея сходные потребности, они различаются между собой размерами яйцекладов, которые у М. greenei Вr. в среднем составляют 4 см, у М. macrurus Br. — 8, у M. atrata Scop. — 11,5 см. Это позволяет разным видам откладывать яйца в личинок, расположенных на различной глубине в древесине заселённых ими деревьев.
Принцип конкурентного исключения можно использовать для поиска возможных различий между сосуществующими близкими видами и оценить при этом действие различных условий на взаимоотношения между ними. Например, вытеснение суринамского мукоеда (Oryzaephilus surinamensis L.) малым мучным хрущаком (Tribolium confusum Duv.) возможно предотвратить, поместив в муку тонкие стеклянные трубочки. Используемые как убежища сравнительно мелкими личинками мукоеда, они сразу восстанавливают равновесие между конкурирующими видами.
При сильной межвидовой конкуренции ареалы конкурирующих видов сокращаются до оптимальных границ и остаются относительно неизменными. Если внутривидовые механизмы рассредоточения популяций превышают давление конкуренции, то ареал расширяется за пределы оптимума; при этом начинают действовать иные лимитирующие факторы.
Принцип конкурентного исключения позволяет очертить границы экологической ниши вида. Концепция экологической ниши — одна из важнейших в современной экологии. По образному выражению Ю. Одума, она представляет собой «профессию вида», а местообитание — его «адрес». Однако среди множества определений этого понятия нет ни одного общепринятого. По одному из них экологическая ниша — это диапазон условий, при которых существуют и воспроизводятся популяции, по другому — экологическая ниша равна общей сумме адаптации. Очевидно, сколько-нибудь полное описание ниши можно представить как бесконечный ряд частных характеристик среды и свойств обитающих в ней организмов.
Приведённые примеры конкурентного исключения выявляют лишь некоторые, наиболее важные параметры, гарантирующие сосуществование конкурентов. В простейшем случае, ограничившись зависимостью организмов от какого-либо одного меняющегося фактора среды, можно дифференцировать экологические ниши разных видов. Основываясь, в частности, на законе толерантности Шелфорда и полагая, что приспособленность популяций соответствует их успеху в воспроизводстве, можно выделить разные варианты взаимодействия двух видов (рис. 186).

Рисунок 213. Сеноеды (по Гилярову, 1969)
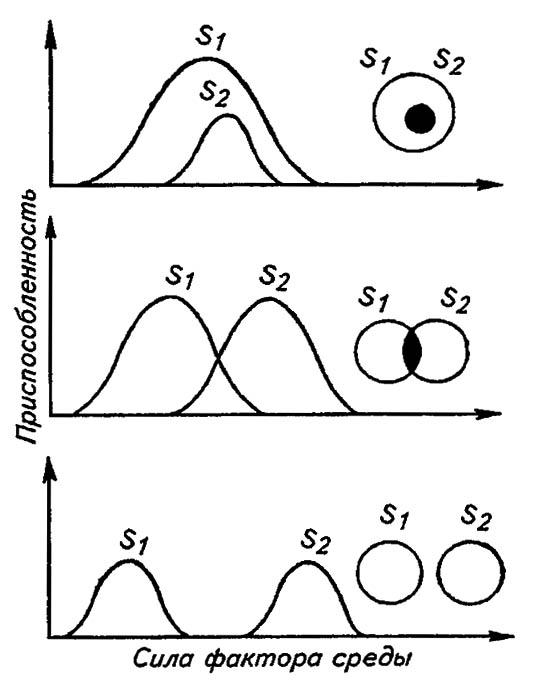
Рисунок 186. Примеры возможных взаимодействий экологических ниш двух (S1 и S2) видов (по Пиаике, 1981)
В случае, когда ниши полностью разобщены, населяющие их виды не конкурируют друг с другом. Если они вообще не вступают в конкуренцию с другими организмами, то их ниши близки к потенциально возможным в данной ситуации. Например, первые наземные обитатели оказались в своеобразном экологическом вакууме. Освободившись от конкуренции с водными организмами, они быстро заселили все возможные местообитания и, приобретя все возможные «профессии», распространились по всем доступным экологическим нишам. Насекомые, обретя крылья, первыми из наземных существ вырвались в новые адаптивные зоны и лишь впоследствии рассредоточились по более тесным экологическим нишам, ограничиваемым воздействиями конкурентов.
В случае, когда экологические ниши частично совпадают, сосуществование видов становится возможным благодаря специфичности приспособлений, например, если имеются убежища, недоступные для более активного конкурента.
Наконец, в случае, когда ниша одного вида включает в себя нишу второго или они полностью перекрываются, произойдёт конкурентное исключение или доминирующий конкурент оттеснит своего соперника на периферию зоны приспособленности. В этом случае реальная экологическая ниша слабого конкурента предельно сузится, заняв лишь некоторую часть потенциально возможной ниши.
К наиболее очевидным способам разобщения экологических ниш относятся: их пространственная изоляция, пищевая избирательность, различия в требованиях к микроклимату населяющих ниши организмов, а также дифференциация сезонной и суточной активности организмов.
Например, развивающиеся синхронно личинки стрекоз Coenagrion resolutum Ken. и Enalagma boreale Charp. избегают конкуренции, заселяя либо глубокие открытые водоёмы, либо заросшие и мелкие. Случайно завезённый в 1900 году в Калифорнию паразит цитрусовой щитовки Aphidius chrysomphagis Zt. распространился со своим хозяином во все цитрусоводческие районы. Впоследствии, в 1948 г., для усиления борьбы с щитовкой из Китая были интродуцированы паразиты близкого вида — A. lingnanensis Kyr., которые, вытеснив первый вид отовсюду, кроме отдельных мест на побережье, оказались малоэффективными в подавлении численности щитовки. Во внутри-континентальных районах, подверженных большим перепадам температуры и влажности, новый вид перестал размножаться из-за холодовой инактивации спермиев, и потому снизилась эффективность защиты цитрусовых культур. Чтобы исправить создавшееся положение, в 1956 и 1957 годах из Пакистана и Индии был интродуцирован третий вид этого рода — A. melinus Br., который, в свою очередь, вытеснив A. lingnanensis Kyr., содействовал успеху биологической борьбы с вредителем.
Сходным образом в США колорадский картофельный жук (Leptinotarsa decemlineata Say.), перешедший с Solarium rostratum на картофель (S. tuberosum) и S. carolinensis, постепенно вытеснил с последнего его потребителя — Leptinotarsa juncta Say. во всех районах, кроме Флориды.
Примером дифференциации ниш по микроклимату является различие в потребностях двух видов крестоцветных блошек, повреждающих капусту: Phyllotreta crucifera F. и Ph. striolata Curt. Первый, светолюбивый, вид сосредоточивается на вершинах листьев, второй, теневыносливый, — у их основания. В результате оба вида существуют на одном растении без конкурентного исключения.
Способности близких видов использовать одни и те же ресурсы, но в разное время выражены у пчёл рода Andrena. Вид A. chylismia L. посещает цветки рано утром, а вид A. rozeni Kby. — в конце дня. Ещё более резкая дифференциация во времени наблюдается у дневных и сумеречных насекомых и насекомых, заселяющих сходные субстраты в разные сроки. Все это позволяет избежать взаимной конкуренции.
Хищничество и паразитизм
При этих формах взаимодействия популяций агрессивности хищников и паразитов противостоят способности их жертв к самозащите и усиленному воспроизводству.
Отличия хищников от паразитов состоят в том, что первые используют свою жертву однократно и умерщвляют её, вторые более «расчётливы»; не заинтересованные в быстрой гибели хозяина, они пользуются им долговременно. Хищники, как правило, крупнее своей добычи; паразиты обычно мельче хозяев и нередко существуют в их теле, получая не только корм, но и защиту. Сопряжённость такого рода взаимодействий особенно наглядна в абстрактной модели «хищник—жертва», разработанной Лоткой и Вольтерра. Здесь повышение численности жертв стимулирует размножение хищников (численная реакция). Вскоре чрезмерное обилие хищников приводит к резкому падению численности жертв, что, в свою очередь, ведёт к вымиранию популяций хищников. Освободившись от хищников, оставшиеся жертвы быстро восстанавливают свою численность, провоцируя последующее размножение сохранившихся хищников. Все это проявляется в периодичности колебаний численности тех и других, и эти колебания в модельных ситуациях продолжаются как угодно долго.
В реальных условиях при резком сокращении числа жертв многие хищники и некоторые паразиты переключаются на другие виды и тем самым избегают последствий собственной прожорливости. Их численность остаётся стабильной. На этом основано утверждение о том, что видовое разнообразие сообществ сопряжено с их стабильностью и всякое нарушение стабильности биоценоза провоцирует выделение доминирующих видов при общем сокращении видового разнообразия. Даже удаление хищника содействует сокращению видового разнообразия сообществ, так как хищник нередко служит стабилизатором численности более активного конкурента. При длительном совместном культивировании, например, комнатной мухи (Musca domestica L.) с паразитической хальцидой Nasonia vitropennis L. после рада колебаний, соответствующих модели хищник — жертва, их численность стабилизируется (рис. 187).
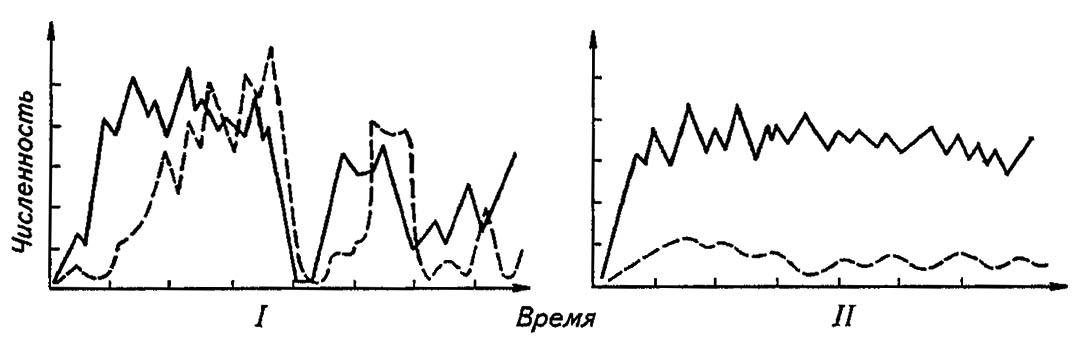
Рисунок 187. Стабилизация численности хищника и жертвы при длительном совместном культивировании (по Одуму, 1975):
сплошная линия — Musca domestica, пунктирная — Nasonia vitropennis; I — исходное состояние; II — состояние стабилизации
Хищничество — обычное явление среди насекомых; к их жертвам, как правило, относятся другие насекомые. Однако крупные тропические богомолы легко справляются с мелкими ящерицами, плотоядные личинки стрекоз ловят мелких рыбок, а некоторые осы, вступая в единоборство с пауками, уносят их в свои гнезда в парализованном состоянии. Даже среди таких типично растительноядных форм, как бабочки, имеется не менее 60 хищных видов.
Значение хищничества кроме показанного выше определяется влияниями на популяции жертв. Особи, избежавшие воздействия хищничества, дают начало потомкам, наследующим способности своих родителей и передающим их, в свою очередь, своим потомкам. Среди популяции хищников также происходит отбор: выживают и оставляют потомство только те из них, которые сумели настигнуть свои жертвы и победить их в борьбе.
Явление паразитизма значительно сложнее, чем хищничество. Существует несколько форм взаимодействия паразита с хозяином; самая распространённая — облигатный паразитизм. К облигатным паразитам относятся пухоеды, вши, блохи и веерокрылые, которые вообще не способны существовать без хозяев, оказывая на их организм большее или меньшее воздействие. Среди насекомых имеются также паразиты временные и факультативные, для которых связь с хозяином эпизодична или необязательна.
Веерокрылые и многие другие паразитические насекомые относятся к энгомофагам, то есть избирают в качестве хозяев других представителей своего класса. Например, поражение пчёл веерокрылыми сопровождается кастрацией, и стерилизованные таким образом особи, оставаясь живыми, развивают внешние признаки противоположного пола. Как энтомофаги особенно примечательны перепончатокрылые. В отличие от типичных паразитов они имеют облик и повадки хищников. Крупные (до 5 см) осы-сколии, или тифии, охотясь на личинок пластинчатоусых жуков, парализуют их точным уколом жала в нервные центры. На обездвиженные таким образом жертвы они откладывают по одному яйцу, и выходящая из него личинка осы начинает пожирать личинку жука. Сначала она съедает жировое тело и другие органы, не трогая нервную и кровеносную системы, поддерживающие жизнь хозяина. Изредка личинка жука оправляется от паралича и обретает активность, но она уже не способна избавиться от паразита.
Парализованные наездником Bracon juglandis L. гусеницы мельничной огнёвки (Ephestia kuhniella Zell.) теряют подвижность, но сохраняют слабые реакции на раздражители. Их метаболизм тормозится, но спинной сосуд продолжает пульсировать долгое время (до 52 дней после нападения), меняя частоту биений в зависимости от температуры.
Необратимый паралич жертвы можно рассматривать как исходный и относительно примитивный способ овладения ею. Более совершенные паразиты вызывают временную иммобилизацию хозяина или откладывают в его тело свои яйца, не ограничивая его активности. Усложнение форм паразитизма самих личинок выражается в переходе их от питания на хозяине (экгопаразитизм) к питанию в полости его тела (эндопаразитизм). Кроме того, секреты, вводимые самкой вместе с яйцами в организм хозяина, несколько притормаживают его развитие и блокируют метаморфоз, что, видимо, выгодно её личинкам. Последние под влиянием секрета развиваются быстрее и, избегая неблагоприятных последствий некротических процессов, покидают хозяина до его гибели.
Торможение и подавление развития хозяина могут исходить не только от самки, но и от паразитирующих личинок. Например, многие наездники-яйцееды, развивающиеся в яйцах других насекомых, разрушают ткани зародыша и используют желток для собственного потребления. При поражении наездником Pimpla turionellae Zt. куколок вощинной моли из яиц паразита выделяются ингибирующие вещества, препятствующие метаморфозу. Позднее, когда из этих яиц выходят личинки, они мигрируют в головной мозг куколки, разрушают его и заканчивают развитие в брюшке. По-видимому, все это служит приспособлением для тех видов, которые, паразитируя в быстро проходящих фазах развития хозяина, искусственно продлевают или прерывают их течение. Однако многие наездники-яйцееды, заражая яйцо, завершают развитие в формирующихся личинках и куколках хозяина, другие (например, заселяющий личинок луковой мухи Hylemyia antiqua Mg. наездник Aphaereta pallipes St.) ускоряют их метаморфоз и окукливание.
Обычно паразит угнетает активность хозяина, но иногда, наоборот, содействует более интенсивному потреблению пищи или провоцирует его к таким действиям, которые выгодны самому паразиту. Так, жуки Pissodespiceae Ill., поражённые Brachistes sp., строят куколочные колыбельки не в глубине дерева, а вблизи его поверхности, что облегчает выход паразитов наружу.
Влияние паразитов на организм хозяина далеко не всегда однозначно. Потенциальные хозяева обладают различными маскировочными приспособлениями и средствами самозащиты. Некоторые из них спасаются бегством или спрыгивают с растений, заметив энтомофага. Другие принимают угрожающие позы, выбрызгивают резко пахнущие или ядовитые секреты либо стремятся сбросить паразита со своего тела.
В связи с тем что покоящиеся фазы развития хозяина лишены средств активной самозащиты, многие энтомофаги, не способные овладеть активным насекомым, специализируются на поиске и заражении его яиц и куколок. Если заражение произошло, то хозяин стремится освободиться от отложенных на его тело яиц, сбрасывая их или повреждая челюстями. Однако нередко паразит прикрепляет яйца в складках покровов, межсегментных бороздах и в тех местах, которые не могут быть очищены придатками тела хозяина. Например, Erromenus sp. прикрепляет яйца к стенкам задней кишки личинок пилильщиков, а Exenterus sp. снабжает свои яйца крючьями, препятствующими их удалению с поверхности тела.
Особый интерес представляют физиологические защитные реакции хозяина, направленные против паразитов, проникших в его тело. Нередко вокруг паразитов образуются соединительнотканные капсулы, препятствующие движению и дыханию. У гусениц мельничной огнёвки эти капсулы формируются уже на третьи сутки после заражения, и заключённый в них паразит вскоре погибает от удушья. Известно много примеров гибели паразитов в теле хозяина без формирования капсул. По-видимому, здесь вступают в действие такие факторы самозащиты, как гемоциты и продуцируемые ими ферменты.
Многие паразиты избегают контакта с гемоцитами, внедряясь, например, в ганглии нервной системы или подавляя их активность. Некоторые специализированные энтомофаги препятствуют образованию капсул, выделяя физиологически активные вещества. Иногда эту роль выполняют секреты, вводимые самкой в тело хозяина вместе с яйцами.
Подавлению защитных сил поражённого организма способствует выбор наименее устойчивых к заражению фаз развития хозяина. В начале онтогенеза, когда форменные элементы гемолимфы представлены лишь прогемоцитами, защитных соединительнотканных капсул не образуется. Поэтому выбор для заражения яиц и личинок младших возрастов гарантирует паразиту относительно спокойное существование.
Суперпаразитизм (перезаражение) — заражение хозяина сразу многими яйцами — содействует ослаблению защитных реакций инкапсуляции, так как на каждое яйцо приходится сравнительно немного гемоцитов. Примером может служить заражение долгоносика Hypera brunneipennis Сар. несколькими яйцами наездника Bathyplectes curculionis Sm. Если одиночные паразиты часто гибнут внутри капсул, то при откладке в тело хозяина, например, четырёх яиц их шансы на выживание соответственно возрастают. Однако при этом между вышедшими из яиц личинками могут возникнуть конкурентные отношения, и эффект перезаражения нейтрализуется из-за внутривидовой конкуренции.
Взаимодействия в системе паразит—хозяин, проявляющиеся в развитии защитных приспособлений и средств их преодоления, отражают сложность и противоречивость эволюционного процесса. Даже близкие виды часто сильно различаются между собой по интенсивности защитных реакций. Например, Hypera postica Schrank. инкапсулирует до 4 % яиц наездника Bathyplectes curculionis Sm., а Н. brunneipennis L. — до 80 %.
Внутривидовую изменчивость реакций и направленный отбор на развитие устойчивости можно проиллюстрировать следующими примерами. Северные популяции репной белянки (Pieris rapae L.) более устойчивы к паразиту Apanteles glomeratus L., чем южные. Характерный пример — акклиматизация наездника Mesoleius tenthredinis Ten., завезённого в Северную Америку для борьбы с лиственничным пилильщиком Pristiphora erichsonii Lep. В 20-х годах нашего столетия он эффективно контролировал численность вредителя, но начиная с 1940 года пилильщик приобрёл высокую устойчивость к заражению. Это потребовало интродукции новых видов и рас М. tenthredinis Ten., способных преодолеть сформировавшиеся физиологические барьеры хозяина.
Наряду с одиночным и групповым паразитизмом, к частному случаю которого относится перезаражение, или суперпаразитизм, среди паразитов-энтомофагов встречается гиперпаразитизм. Этим термином обозначают явление, когда паразит сам становится хозяином для другого паразита, именуемого сверхпаразитом первого порядка. Не исключено заражение и сверхпаразитов, но сверхпаразиты второго порядка встречаются очень редко. Один из примеров — заражение наездником Asecodes albitarsus Walker многоядного сверхпаразита Dibrachys boucheanus Bouch., который, в свою очередь, нередко поражает наездников Apanteles glomeratus L., существующих за счёт гусениц бабочек-белянок.
Одна из форм паразитизма — клептопаразитизм (буквально: воровской паразитизм). Клептопаразит использует активность другого паразита для своих нужд, в частности для облегчения поиска и заражения хозяина. Нередко он откладывает свои яйца через отверстия в покровах ранее заражённого хозяина и устраняет личинок первого паразита в межвидовой конкуренции.
Весьма своеобразный пример отрицательных взаимодействий популяций разных видов представляют собой некоторые виды муравьёв, нападающие на гнезда других видов и уносящие после ожесточённых сражений их личинок и куколок. Воспитывая захваченных особей в собственном муравейнике, они используют их впоследствии на разных работах, не выпуская наружу. «Рабовладельчество» характерно для Formica sanguinea Latr., эксплуатирующих особей F. rufibarbis F., и для муравьёв-амазонок Polyargus rufescens L. Последние возлагают на своих «рабов» все работы по содержанию гнезда и уходу за личинками и даже питаются только с их помощью.
Положительные взаимодействия популяций менее изучены, чем отрицательные. Они наблюдаются обычно в стабильных, ненарушенных биоценозах. Различают несколько форм взаимодействий по сопряжённости существования популяций, по значению для их воспроизводства, но провести резкие границы между этими взаимодействиями не всегда возможно. Некоторые из них сближаются с отрицательными взаимодействиями и в определённых ситуациях переходят в них. Это затрудняет их дифференциацию и изучение.
Мутуализм, или симбиоз, представляет собой взаимовыгодное, часто необходимое сосуществование разных видов. Примером мутуализма может служить симбиоз термитов с населяющими их кишечник жгутиконосцами. Утратив симбионтов, термиты гибнут от голода, не имея собственных ферментов для переваривания клетчатки. Сами симбионты вообще не способны существовать во внешней среде и в организмах других насекомых, кроме некоторых тараканов.
Симбиотические организмы отмечены у многих равнокрылых, у некоторых жуков, чешуекрылых, двукрылых и перепончатокрылых. При этом их передача от поколения к поколению гарантируется трофаллаксисом, поеданием заражённых экскрементов и оболочек яиц, или трансовариальным переносом из клеток жирового тела через стенку яичника в формирующиеся яйца.
Несколько меньшая зависимость проявляется в мутуалистических отношениях тлей и муравьёв. Поедая сахаристые экскременты тлей и защищая их от врагов, муравьи переносят тлей на новые кормовые растения, содействуя расселению вредоносных видов. Так, посещения колоний бобовой тли (Aphisfabae Scop.) муравьями Lasius niger L. стимулируют их воспроизводство. Муравьи «ухаживают» также за листоблошками, червецами и щитовками, получая при этом высококалорийную углеводную пищу.
Такие же взаимоотношения наблюдаются между Formica cinerea Mayr. и гусеницами бабочки-голубянки (Lycaena argus L.), зарывающимися в муравейник перед окукливанием, или между муравьями других видов и мирмекофильными жуками из семейств Paussidae, Pselaphidae, Staphilinidae. Они выделяют секреты, охотно поедаемые муравьями; при этом муравьи защищают их от хищников и переносят с другими членами семьи в новые места при её переселении. Однако стафилины родов Lomechusa и Atemeles, обитая в муравейнике, оказываются жестокими хищниками. «Опьяняя» муравьёв своими специфическими секретами, они быстро уничтожают их личинок и яйца.
Синойкией, или сожительством, называют отношения, полезные для одного вида, но безразличные или необременительные для другого. Многие термитофилы и мирмекофилы, находящие приют в термитниках и муравейниках, обретают в них надёжную защиту от врагов и неблагоприятных климатических условий. К ним относятся клещи и ногохвостки, двукрылые, жуки и другие насекомые-сапрофаги, питающиеся разлагающимися растительными остатками и мицелием грибов.
Своеобразное проявление синойкии — форезия, то есть использование других видов для расселения. Например, многие мучные клещи используют насекомых как средство передвижения, а ногохвостки избирают для транспорта грызунов, не причиняя им никаких неудобств.
Комменсализм, или нахлебничество, — это использование одним видом пищевых запасов другого вида, имеющихся в избытке. Не достигающее уровня конкуренции и не ощущаемое партнером, это взаимодействие сближается, с одной стороны, с синойкией, а с другой — с паразитизмом или хищничеством. Например, муха Termitoxenia получает от термитов ту же пищу, которой они делятся со своими сородичами. Постоянно обитая в термитнике, эта муха утратила ненужные здесь крылья и стала гермафродитом, не нуждающимся в половом партнёре.
Крошечные муравьи Formicoxenus, поселяясь с рыжими лесными муравьями (Formica rufa L.), используют моменты, когда один из них кормит другого. При этом они выхватывают небольшие кусочки пищи у своих более крупных хозяев, которые их не замечают, а если заметят, то не прогоняют.
Примеры взаимовыгодного сотрудничества насекомых разных видов выражены при освоении ими трупов и экскрементов, при ослаблении растений, становящихся доступными для вторичных вредителей, при заселении пожарищ и агроценозов. Жуки-могильщики и мертвоеды, обнаруживая падаль, облегчают мухам и жукам-карапузикам доступ к более мягким тканям, а жукам-кожеедам — к жёстким сухожилиям. При этом они сами получают возможность более длительного питания, как и жуки-навозники, «сотрудничающие» с личинками мух на помете животных. Можно полагать, что положительные взаимодействия популяций обеспечивают более полное и экономное освоение среды. Известно, что среди насекомых, населяющих агроценозы, преобладают отрицательные взаимодействия между видами, но постепенно, по мере стабилизации биоценозов, они уступают место положительным.
Насекомые и растения
Взаимодействия насекомых с растениями оформились на самых ранних этапах эволюции и в дальнейшем совершенствовались параллельно. Растения развивали средства при-
влечения насекомых-опылителей и устойчивость к наносимым ими повреждениям; в свою очередь, насекомые совершенствовались как фитофаги. Важнейший результат этих взаимодействий — современное разнообразие покрытосеменных и обилие связанных с ними насекомых, сравнимых по характеру связей с комменсалами, симбионтами и паразитами и лишь в редких случаях — с хищниками. В целом эти взаимоотношения следует признать положительными и весьма важными не только для отдельных биоценозов, но и для всей биосферы.
Подавляющее большинство (до 80 %) высших цветковых растений опыляется при участии насекомых, другие вторично приспособились к опылению ветром и самоопылению. Трудно переоценить значение этого факта — все многообразие цветов предназначено для насекомых.
В свою очередь, насекомые приобрели замечательные приспособления для сбора пыльцы и нектара, особенно характерные у пчелиных и бабочек-бражников. Погружая длинные хоботки в глубину нектарника, они касаются тычинок и затем переносят на другие цветки приставшую к телу пыльцу. Нектарники имеют небольшое количество нектара, привлекающего насекомое, но недостаточное для его насыщения. Это стимулирует насекомых к посещению многих цветков и обеспечивает перекрёстное опыление. Примечательно, что завезённый в Австралию клевер ползучий не давал семян до тех пор, пока не были завезены шмели — его специфические опылители.
Взаимоотношения растений и насекомых нередко достигают предела специализации. Например, люцерну опыляют одиночные пчелы, цветки какао — мелкие мокрецы, инжир — перепончатокрылые Blastophaga psenes L. Бескрылые самцы этого вида оплодотворяют крылатых самок в полом цветоложе короткопестичных цветков неплодоносящих деревьев — каприфиг. Перелетая на длиннопестичные цветки плодоносящих деревьев (фиги), самки переносят на них пыльцу с каприфиг, но так как они имеют короткий яйцеклад, то не могут здесь же отложить яйца. Они возвращаются на каприфиги, перезимовывают в их цветках и спариваются с самцами.
Интересен следующий пример. Известный эволюционист и путешественник А. Уоллес обнаружил на острове Мадагаскар орхидею с глубоким нектарником, недоступным для описанных к тому времени насекомых. Предположив, что её опыление происходит при участии ещё не открытого вида бражников Macroglossa predicta W. (лат. predicta — предсказанный), он вскоре получил подтверждение своей догадке, основанной на существовании глубокой сопряжённости растений и насекомых. Однако некоторые лишённые нектарников орхидеи привлекают опылителей другими способами. Одни из них похожи на цветки видов, производящих нектар, другие — на самок бабочек или на хозяев насекомых-опылителей.
Многие посещающие цветки насекомые не относятся к опылителям или не имеют приспособлений для переноса пыльцы. Таковы, например, муравьи, некоторые жуки, осы и мухи. Однако есть растения, имеющие своеобразные цветки-ловушки, которые принуждают случайных посетителей к выполнению функций опыления. Эти цветки задерживают насекомых, и тычинки размазывают по их телу пыльцу.
Насекомые избирают растения для откладки яиц и для питания вылупляющихся из них личинок. Один из аспектов взаимоотношений с растениями определяется ролью насекомых как фитофагов, объедающих листву и другие вегетативные органы, а часто и цветки, бутоны, формирующиеся плоды. Велика вредоносность насекомых на посевах культурных растений в агроценозах.
Остановимся на значении вторичных метаболитов растений, ядовитых для многих насекомых, но привлекательных для избранных. Содействуя поиску и выбору производящих эти соединения растений, вторичные метаболиты снижают напряжённость конкуренции с другими, не приспособившимися к ним фитофагами и, кроме того, придают ядовитость своим специфическим потребителям. Защищая их таким образом от конкурентов и хищников, а также благодаря выраженному антибиотическому действию и от патогенов вторичные метаболиты растений обеспечивают дифференциацию пищевых запросов (и экологических ниш) насекомых, их избирательность и специфичность. Примечательно, что эти соединения распределяются неравномерно. Так, в цветках зверобоя (Hypericum perforatum L.) концентрация ядовитого гиперицина в 10 раз выше, чем в стеблях и листьях. Естественно, что многие насекомые избегают повреждать генеративные органы этого растения, привлекательные лишь для некоторых опылителей.
Специализация насекомых как фитофагов проявляется в наносимых ими повреждениях (рис. 188), детальная классификация которых составляет предмет сельскохозяйственной энтомологии. Наряду с повреждениями, не требующими предварительной подготовки кормового субстрата, некоторые насекомые сначала скручивают листья в узкие трубочки (тли, жуки-трубковерты и др.), внутри которых они спокойно высасывают соки растения. Другие вырезают из листа маленькие пластинки и потребляют их после предварительного силосования в земляных норках (жуки-кравчики). Галлообразователи (галлицы, орехотворки, некоторые тли и др.), приступая к питанию, стимулируют разрастание тканей в виде бесформенных опухолей, или галлов, имеющих вид орешков, бородавок и тому подобного. Нередко галлообразователи остаются в полости растения, обеспечивая тем самым себе не только питание, но и защиту.

Рисунок 188. Различные типы повреждений, вызываемые насекомыми (по разным авторам)
Фитофаги причиняют огромный вред, однако нет ни одного вида растений, уничтоженных насекомыми полностью. Растения существуют, несмотря на исключительную прожорливость фитофагов.
Защитные реакции и приспособления растений многообразны: структура и прочность покровов, ядовитость, шипы и колючки ограничивают агрессивность фитофагов. Способность растений к регенерации содействует быстрому восстановлению объеденных листьев, ветвей и корней. К одному из наиболее ярких примеров устойчивости растений к вредителям, основанной на особенностях строения, относятся выведенные В. С. Пустовойтом «панцирные» сорта подсолнечника. Формирование углеродистого слоя между пробковой тканью и склеренхимой семян препятствует питанию подсолнечниковой огнёвки (Homoeosoma nebulellum Hb.), повреждающей все другие сорта. Важнейшие защитные реакции хвойных растений выражаются в обильном выделении смолы из повреждений или в утолщении пробкового слоя, индуцированном нападением тлей-хермесов.
Таким образом, насекомые необходимы растениям как опылители, но вместе с тем это группы наиболее массовых фитофагов. Причиняемый ими вред не ограничивается питанием на растениях и откладкой яиц в их ткани и органы. Некоторые насекомые известны как переносчики болезней растений. Однако взаимодействия насекомых с растениями в целом, сформировавшиеся как результат длительной сопряжённой эволюции, можно оценить как гармоничные и прогрессивные.

