9.5.2. Fungus farming by leaf-cutter ants
The subterranean ant nests of the genus Atta (15 species) and the rather smaller colonies of Acromyrmex (24 species) are amongst the major earthen constructions in neotropical rainforest. Calculations suggest that the largest nests of Atta species involve excavation of some 40 tonnes of soil. Both these genera are members of a tribe of myrmecine ants, the Attini, in which the larvae have an obligate dependence on symbiotic fungi for food. Other genera of Attini have monomorphic workers (of a single morphology) and cultivate fungi on dead vegetable matter, insect feces (including their own and, for example, caterpillar “frass”), flowers, and fruit. In contrast, Atta and Acromyrmex, the more derived genera of Attini, have polymorphic workers of several different kinds or castes (section 12.2.3) that exhibit an elaborate range of behaviors including cutting living plant tissues, hence the name “leaf-cutter ants”. In Atta, the largest worker ants excise sections of live vegetation with their mandibles (Fig. 9.6a) and transport the pieces to the nest (Fig. 9.6b). During these processes, the working ant has its mandibles full, and may be the target of attack by a particular parasitic phorid fly (illustrated in the top right of Fig. 9.6a). The smallest worker is recruited as a defender, and is carried on the leaf fragment.
When the material reaches the nest, other individuals lick any waxy cuticle from the leaves and macerate the plant tissue with their mandibles. The mash is then inoculated with a fecal cocktail of enzymes from the hindgut. This initiates digestion of the fresh plant material, which acts as an incubation medium for a fungus, known only from these “fungus gardens” of leaf-cutter ants. Another specialized group of workers tends the gardens by inoculating new substrate with fungal hyphae and removing other species of undesirable fungi in order to maintain a monoculture. Control of alien fungi and bacteria is facilitated by pH regulation (4.5–5.0) and by antibiotics, including those produced by mutualistic Streptomyces bacteria associated with ant cuticle. In darkness, and at optimal humidity and a temperature close to 25°C, the cultivated fungal mycelia produce nutritive hyphal bodies called gongylidia. These are not sporophores, and appear to have no function other than to provide food for ants in a mutualistic relationship in which the fungus gains access to the controlled environment. Gongylidia are manipulated easily by the ants, providing food for adults, and are the exclusive food eaten by larval attine ants. Digestion of fungi requires specialized enzymes, which include chitinases produced by ants from their labial glands.
A single origin of fungus domestication might be expected given the vertical transfer of fungi by transport in the mouth of the founding gyne (new queen) and regurgitation at the new site. However, molecular phylogenetic studies of the fungi show domestication from free-living stocks has taken place several times, although the ancestral symbiosis is at least 50 million years old. All but one domesticate belongs to the Basidiomycetes of the tribe Leucocoprini in the family Lepiotaceae, propagated as a mycelium or occasionally as a unicellular yeast. Although each attine nest has a single species of fungus, amongst different nests of a single species a range of fungus species are tended. Obviously, some ant species can change their fungus when a new nest is constructed, perhaps when colony foundation is by more than one queen (pleiometrosis). Lateral (horizontal) transfer was observed when a Central American Atta species introduced to Florida rapidly adopted the local attinetended fungus for its gardens.
Leaf-cutter ants dominate the ecosystems in which they occur; some grassland Atta species consume as much vegetation per hectare as domestic cattle, and certain rainforest species are estimated to cause up to 80% of all leaf damage and to consume up to 17% of all leaf production. The system is an effective converter of plant cellulose to usable carbohydrate, with at least 45% of the original cellulose of fresh leaves converted by the time the spent substrate is ejected into a dung store as refuse from the fungus garden. However, fungal gongylidia contribute only a modest fraction of the metabolic energy of the ants, because about 95% of the respiratory requirements of the colony is provided by adults feeding on plant sap from chewed leaf fragments.
Leaf-cutter ants may be termed highly polyphagous, as studies have shown them to utilize between 50 and 70% of all neotropical rainforest plant species. How- ever, as the adults feed on the sap of fewer species, and the larvae are monophagous on fungus, the term polyphagy strictly may be incorrect. The key to the relationship is the ability of the worker ants to harvest from a wide variety of sources, and the cultivated fungus to grow on a wide range of hosts. Coarse texture and latex production by leaves can discourage attines, and chemical defenses may play a role in deterrence. However, leaf-cutter ants have adopted a strategy to evade plant defensive chemicals that act on the digestive system: they use the fungus to digest the plant tissue. The ants and fungus co-operate to break down plant defenses, with the ants removing protective leaf waxes that deter fungi, and the fungi in turn producing carbohydrates from cellulose indigestible to the ants.
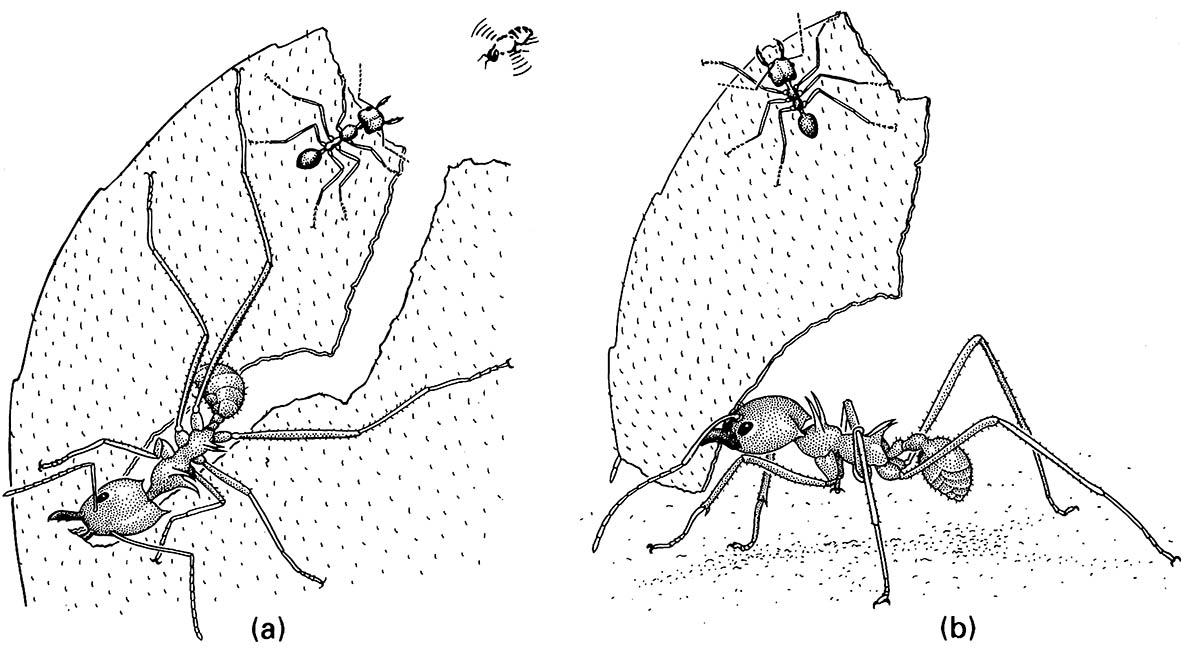
(a) A medium-sized worker, called a media, cuts a leaf with its serrated mandibles while a minor worker guards the media from a parasitic phorid fly (Apocephalus) that lays its eggs on living ants. (b) A guarding minor hitchhikes on a leaf fragment carried by a media. (After Eibl-Eibesfeldt & Eibl-Eibesfeldt 1967)

