16.5.1. Arthropod natural enemies
Entomophagous arthropods may be predatory or parasitic. Most predators are either other insects or arachnids, particularly spiders (order Araneae) and mites (Acarina, also called Acari). Predatory mites are important in regulating populations of phytophagous mites, including the pestiferous spider mites (Tetranychidae). Some mites that parasitize immature and adult insects or feed on insect eggs are potentially useful control agents for certain scale insects, grasshoppers, and stored-product pests. Spiders are diverse and efficient predators with a much greater impact on insect populations than mites, particularly in tropical ecosystems. The role of spiders may be enhanced in IPM by preservation of existing populations or habitat manipulation for their benefit, but their lack of feeding specificity is restrictive. Predatory beetles (Coleoptera: notably Coccinellidae and Carabidae) and lacewings (Neuroptera: Chrysopidae and Hemerobiidae) have been used successfully in biological control of agricultural pests, but many predatory species are polyphagous and inappropriate for targeting particular pest insects. Entomophagous insect predators may feed on several or all stages (from egg to adult) of their prey and each predator usually consumes several individual prey organisms during its life, with the predaceous habit often characterizing both immature and adult instars. The biology of predatory insects is discussed in Chapter 13 from the perspective of the predator.
The other major type of entomophagous insect is parasitic as a larva and free-living as an adult. The larva develops either as an endoparasite within its insect host or externally as an ectoparasite. In both cases the host is consumed and killed by the time that the fully fed larva pupates in or near the remains of the host. Such insects, called parasitoids, all are holometabolous insects and most are wasps (Hymenoptera: especially superfamilies Chalcidoidea, Ichneumonoidea, and Platygas- teroidea) or flies (Diptera: especially the Tachinidae). The Chalcidoidea contains 20 families and perhaps 100,000–500,000 species (mostly undescribed), of which most are parasitoids, including egg parasitoids such as the Mymaridae and Trichogrammatidae (Fig. 16.3), and the speciose ecto- and endoparasitic Aphelinidae and Encyrtidae, which are biological control agents of aphids, mealybugs (Box 16.4), other scale insects, and whiteflies. The Ichneumonoidea includes two speciose families, the Braconidae and Ichneumonidae, which contain numerous parasitoids mostly feeding on insects and often exhibiting quite narrow host-specificity. The Platygasteroidea contains the Platygasteridae, which are parasitic on insect eggs and larvae, and the Scelionidae, which parasitize the eggs of insects and spiders. Parasitoids from many of these wasp groups have been utilized for biological control, whereas within the Diptera only the tachinids are commonly used as biological control agents.
Parasitoids often are parasitized themselves by secondary parasitoids, called hyperparasitoids (section 13.3.1), which may reduce the effectiveness of the primary parasitoid in controlling the primary host — the pest insect. In classical biological control, usually great care is taken specifically to exclude the natural hyperparasitoids of primary parasitoids, and also the parasitoids and specialized predators of other introduced exotic natural enemies. However, some highly efficient natural enemies, especially certain predatory coccinellids, sometimes eliminate their food organisms so effectively that their own populations die out, with subsequent uncontrolled resurgence of the pest. In such cases, limited biological control of the pest’s natural enemies may be warranted. More commonly, exotic parasitoids that are imported free of their natural hyperparasitoids are utilized by indigenous hyperparasitoids in the new habitat, with varying detrimental effects on the biological control system. Little can be done to solve this latter problem, except to test the host-switching abilities of some indigenous hyper-parasitoids prior to introductions of the natural enemies. Of course, the same problem applies to introduced predators, which may become subject to parasitization and predation by indigenous insects in the new area. Such hazards of classical biological control systems result from the complexities of food webs, which can be unpredictable and difficult to test in advance of introductions.
Some positive management steps can facilitate long-term biological control. For example, there is clear evidence that providing a stable, structurally and floristically diverse habitat near or within a crop can foster the numbers and effectiveness of predators and parasitoids. Habitat stability is naturally higher in perennial systems (e.g. forests, orchards, and ornamental gardens) than in annual or seasonal crops (especially monocultures), because of differences in the duration of the crop. In unstable systems, the permanent provision or maintenance of ground cover, hedgerows, or strips or patches of cultivated or remnant native vegetation enable natural enemies to survive unfavorable periods, such as winter or harvest time, and then reinvade the next crop. Shelter from climatic extremes, particularly during winter in temperate areas, and alternative food resources (when the pest insects are unavailable) are essential to the continuity of predator and parasitoid populations. In particular, the free-living adults of parasitoids generally require different food sources from their larvae, such as nectar and/or pollen from flowering plants. Thus, appropriate cultivation practices can contribute significant benefits to biological control. Diversification of agroecosystems also can provide refuges for pests, but densities are likely to be low, with damage only significant for crops with low EILs. For these crops, biological control must be integrated with other methods of IPM.
Pest insects must contend with predators and parasitoids, but also with competitors. Competitive interactions appear to have little regulative influence on most phytophagous insects, but may be important for species that utilize spatially or temporally restricted resources, such as rare or dispersed prey/host organ- isms, dung, or animal carcasses. Interspecific competition can occur within a guild of parasitoids or predators, particularly for generalist feeders and facultative hyperparasitoids, and may inhibit biological control agents.
Biological control using natural enemies is particularly successful within the confines of greenhouses (glasshouses) or within certain crops. The commercial use of inundative and seasonal inoculative releases of natural enemies is common in many greenhouses, orchards, and fields in Europe and the USA. In Europe, more than 80 species of natural enemies are available commercially, with the most commonly sold arthropods being various species of parasitoid wasps (including Aphidius, Encarsia, Leptomastix, and Trichogramma spp.), predatory insects (especially coccinellid beetles such as Cryptolaemus montrouzieri and Hippodamia convergens, and mirid (Macrolophus) and anthocorid (Orius spp.) bugs), and predatory mites (Amblyseius and Hypoaspis spp.).
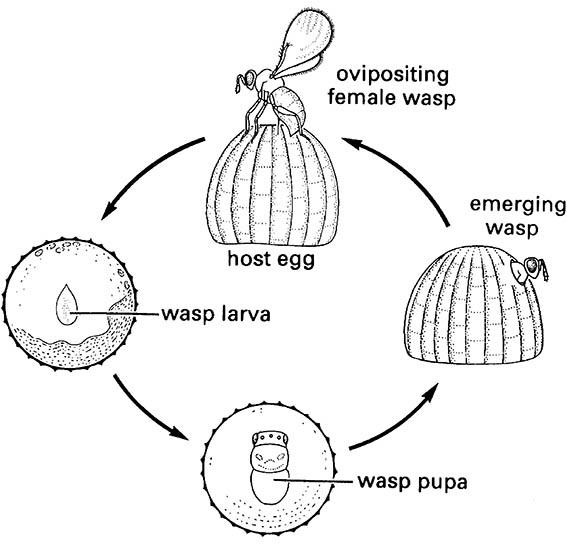
A tiny female wasp of a Trichogramma species (Hymenoptera: Trichogrammatidae) oviposits into a lepidopteran egg; the wasp larva develops within the host egg, pupates, and emerges as an adult, often with the full life cycle taking only one week. (After van den Bosch & Hagen 1966)

