Грудной отдел и его придатки
Сосредоточение локомоторных функций и сопутствующее развитие мышц крыльев и ног привели к обособлению и консолидации грудного отдела насекомых. Слагающие его сегменты, именуемые переднегрудью (prothorax), среднегрудью (mesothorax) и заднегрудью (metathorax), для прочности укреплены дополнительными швами и элементами внутреннего скелета (рис. 26).
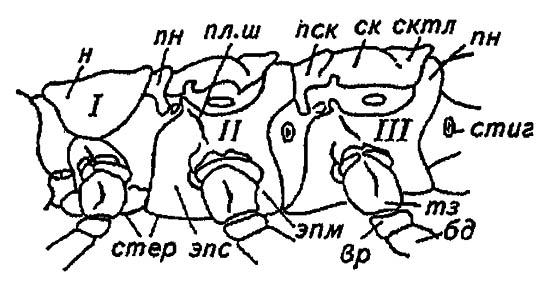
Рисунок 26. Сегменты грудного отдела насекомых (по Шванвичу, 1949):
I — переднегрудь; II — среднегрудь; III — заднегрудь; н — нотум; пн — постнотум; пск, ск, сктл — прескутум, скутум и скутеллум; эпм, эпс — эпимеры и эпистерны плейритов; стер — стерниты; пл. ш — плейральный шов; тз — тазик; вр — вертлуг; бд — бедро; стиг — стигмы
Особенно сильно развились средне- и заднегрудь, на которых у подавляющего большинства имеются крылья. В меньшей степени изменилась лишённая крыльев переднегрудь.
Конструктивной основой грудного сегмента служат связанные друг с другом склериты: спинки — тергит (tergum), груди — стернит (sternum) и боковых стенок — плейрит (pleuron), которые подразделяются на более мелкие элементы внешнего скелета.
Однако прежде чем перейти к их описанию, напомним о несоответствии вторичной сегментации насекомых первичной сегментации кольчатых червей (см. рис. 4).
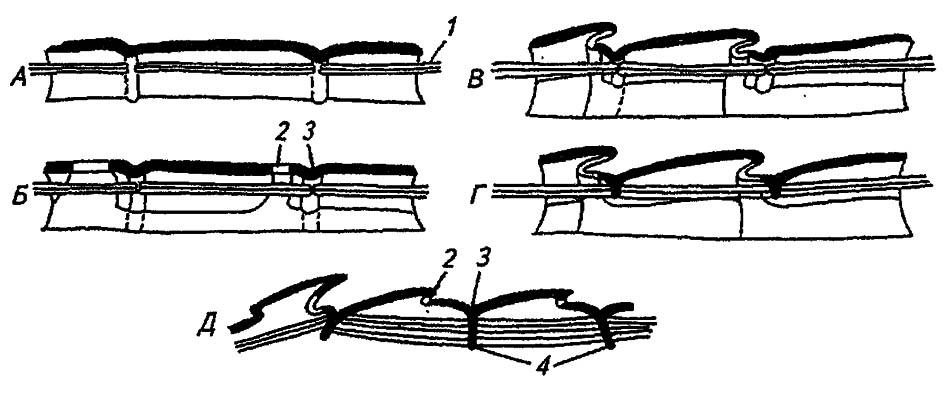
Рисунок 4. Схема вторичной сегментации туловища насекомого (по Шванвичу, 1949, с изменениями):
А — исходное состояние (покровы тела мягкие); Б — склеротизация покровов и формирование вторичных межсегментных борозд; В — телескопическое сдвигание сегментов; Г — образование фрагм; Д — усиление фрагм в крылоносных сегментах с особенно мощной мускулатурой; 1 — продольные спинные мышцы; 2 — вторичные межсегментные борозды; 3 — первичные межсегментные борозды; 4 — фрагмы
Формирующееся в пределах истинного сегмента вторичное сочленение отделяет у заднего края тергита узкий постнотум (postnotum), который соединяется гибкой мембраной с его основной частью — нотумом (notum). Последний для противодействия деформации укреплён парой парапсоидальных швов и непарным V-образным швом, образующими неполную крестовину, препятствующую изгибам тергита (рис. 27).
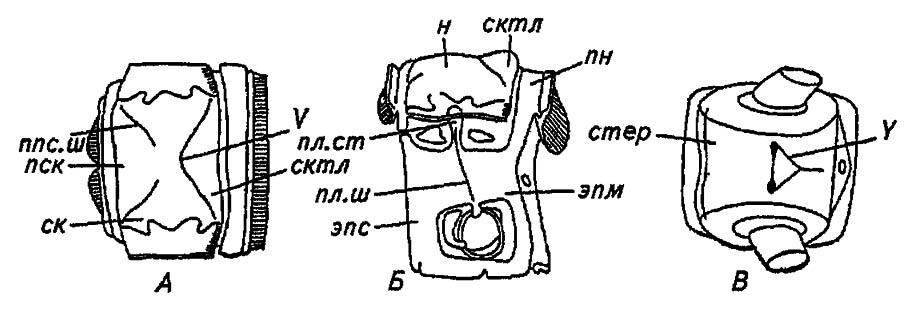
Рисунок 27. Грудной сегмент насекомого в трёх проекциях (по Romoser, 1981):
А — сверху; Б — сбоку; В — снизу; н — нотум; пн — постнотум; ппс. ш — парапсоидальные швы; V — V-образный шов; пск — прескутум; ск — скутум; сктл — скутеллум; пл. ш — плейральный шов; пл. ст — плейральный столбик; Y — Y-образный шов
Её передние ветви ограничивают поле прескутума (praescutum), или предщит, задние ветви — щитик, или скутеллум (scutellum), а пространство, заключённое между ними, называется щитом, или скутумом (scutum).
Сходным образом обеспечивается жёсткость стернита. Расположенные на его поверхности Y-образный шов и стернальная складка, концы которой обозначены глубокими впячиваниями покровов, ограничивают продольные и косые деформации склерита.
Переходя к описанию плейрита, следует указать на разногласия по поводу его природы и происхождения. Вполне развитый в сегментах грудного отдела насекомых, и в особенности в сегментах, несущих крылья, плейрит, видимо, образовался за счёт основного членика — ноги, так называемой субкоксы. В соответствии с субкоксальной теорией, выдвинутой Р. Е. Снодграссом, субкокса, входя в состав стенки тела, образует три дуговидных склерита: анаплейрит и трохантинную пластинку, лежащие над сочленением ноги, и стерноплейрит, расположенный под этим сочленением (рис. 28).
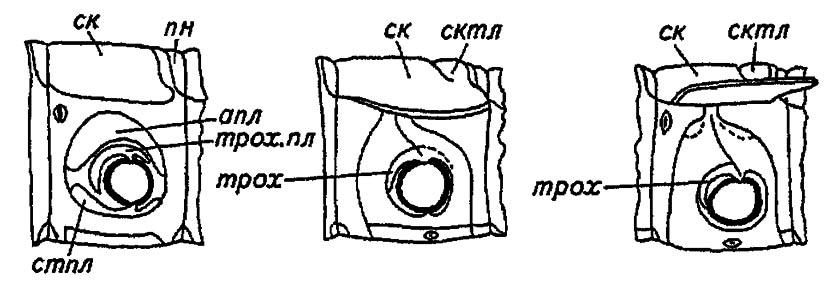
Рисунок 28. Последовательные стадии формирования плейрита насекомых в соответствии с субкоксалыюй теорией происхождения плейрита (по Шванвичу, 1949):
ск — скутум; сктл — скутеллум; пн — постнотум; апл — анаплейрит; стпл — стерноплейрит; трох — трохантин; трлх. пл — трохантинная пластинка
Данное состояние сохраняется у некоторых первичнобескрылых насекомых, тогда как у более совершенных веснянок и термитов стерноплейрит сливается со стернитом, а задняя часть трохантинной пластинки наряду с анаплейритом преобразуется в плейрит. Передняя часть трохантинной пластинки обособляется в виде склерита — трохантина, несущего мыщелок сочленения с конечностью.
У наиболее совершенных насекомых, в частности у перепончатокрылых, трохантин исчезает, и плейральная область становится монолитной. В противовес приведённой трактовке можно сослаться на суждение Э. Г. Беккера, согласно которому многочисленные мелкие склериты плейральной области многоножек, сливаясь друг с другом, послужили основой для формирования тазика и вертлуга конечностей насекомых. Иными словами, не плейрит образовался из конечности, а, наоборот, конечность — из склеритов плейральной области.
Не отрицая ценности обеих гипотез, мы считаем, что развитие плейрита связано с развитием крыльев, чему соответствуют следующие элементы его структуры: эпистерна (episternum) и эпимера (epimeron), разделённые плейральным швом (см. рис. 27), конец которого — плейральный столбик (columella) — обеспечивает опору для движения крыла. На нем, как на оси или стержне весов, раскачивается двуплечий рычаг крыловой пластинки (рис. 29).
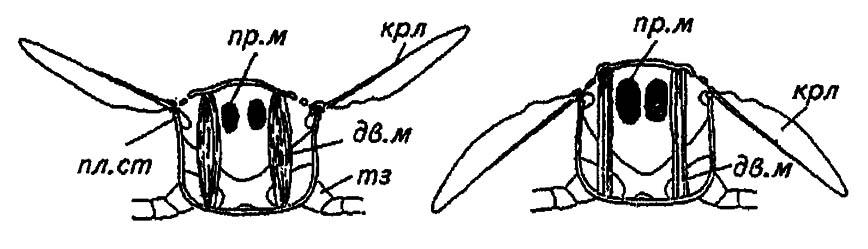
Рисунок 29. Схема строения крылоносного сегмента насекомых (по Шванвичу, 1949):
крл — крылья; пр. м — продольные мышцы; дв. м — дорзовектральные мышцы; тз — тазик; пл. ст — плейральный столбик
Таким образом, плейрит, исключая область сочленения с конечностью, сливается со стернитом в монолитную и прочную капсулу. Вместе с тем сочленение тергита и плейрита сохраняет подвижность: оно представлено гибкой мембраной, которая продолжается жёсткой пластинкой крыла, однако в задней части сегмента отчленённый от него постнотум сливается с эпимерами, что сообщает сегменту в целом дополнительную прочность. При этом каждый крылоносный сегмент (средне- и заднегрудь) приобретает облик чаши или короба, прикрытого сверху крышкой тергита. Сокращения мощных крыловых мышц попеременно притягивают тергит вниз и вместе с ним основания крыльев или, деформируя стенки стерноплейрального комплекса, содействуют выгибанию тергита вверх и наружу, а вместе с ним и движению основания крыльев. Расположенные на плейральном столбике крыловые пластинки соответственно поднимаются вверх или опускаются вниз (см. рис. 29), и эти движения обеспечивают полет насекомых.
Крыловые мышцы подавляющего большинства насекомых (исключая стрекоз) представляют собой мышцы непрямого действия. Движения крыльев обеспечиваются ими через деформации грудных сегментов, и лишь тонкие сухожилия мелких маневрирующих мускулов, выходя из стенки тела и непосредственно причленяясь к склеритам крыла, вызывают изменения его наклона (угла атаки) или складывание вдоль тела. Все эти движения требуют особой прочности стерноплейральной чаши сегмента, которая наряду со швами и бороздами укрепляется элементами внутреннего скелета (см. рис. 5).
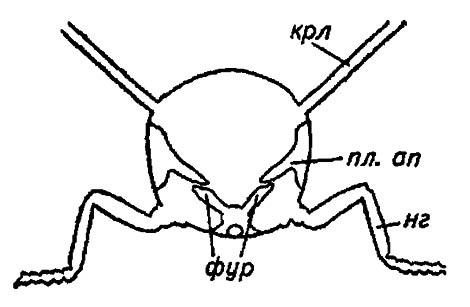
Рисунок 5. Внутренний скелет трудного сегмента насекомого:
крл — крылья; нг — ноги; фур — фурка; пл. ап — плейральные апофизы
Впячивания концов Y-образного шва стернита продолжаются в глубь сегмента в виде фурки (furca) — своеобразной вилочки, свободным концам которой противопоставляются плейральные апофизы (apophysis pleuralis) — выросты плейрального шва.
Приобретение крыльев и способность к полёту открыли перед древними насекомыми ещё никем не освоенную воздушную среду. Лишь много позднее появились громоздкие летающие ящеры, птицы и рукокрылые млекопитающие.
Представляя собой боковые выросты стенки тела, крылья насекомых не гомологичны их конечностям. Они образованы двумя тонкими мембранами, натянутыми на каркас жилок, в которые проникают трахеи, нервы и полостная жидкость. Наиболее крупные жилки хорошо летающих насекомых сосредоточены у переднего края крыла, придавая ему необходимые аэродинамические свойства и жёсткость. У заднего края крыла жилки расположены менее густо и не препятствуют его волнообразным изгибаниям, гасящим стекающие с крыловых плоскостей турбулентные завихрения потоков воздуха при полете. Этим же целям, по-видимому, служат волоски и щетинки, расположенные по заднему краю крыла у многих насекомых.
Не останавливаясь на всем разнообразии крыльев и типов расположения жилок, рассмотрим общий план, или схему, жилкования и наиболее общие тенденции эволюционных преобразований.
Передняя и обычно наиболее толстая костальная жижа (costa) достигает вершины крыла, а иногда распространяется и далее, на его задний край, становясь все более тонкой (рис. 30).
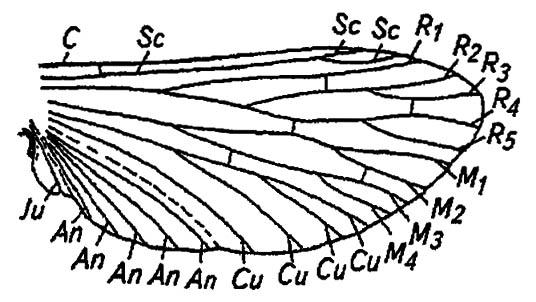
Рисунок 30. Схема жилкования крыла насекомого (по Бей-Биенко, 1966):
C — костальная жилка; Sc — субкостальная жилка; R1 — радиальная жилка; R2–R5 — жилки сектора радиуса; M1 — медиальная жилка; М2–М4 — жилки сектора медианы; Cu, An, Ju — кубитальные, анальные, югальные жилки
За ней расположена субкостальная жилка (subcosta), связанная с костой поперечной плечевой жилкой (humeralis). Нередко у места впадения субкосты в косту формируется плотный кутикулярный противовес — крыловой глазок (pterostigma). Это образование снимает вредную вибрацию — флаттер крыловых плоскостей при полете. Далее находится радиальная жилка (radius) с отходящими от неё жилками сектора (radii sector). Выходя на край крыла у его вершины, ветвящиеся жилки сектора радиуса нумеруются спереди назад, как показано на рисунке. Близ середины крыла, непосредственно за сектором радиуса, расположены ветвящиеся медиальные жилки: первая из них — медиана (mediana), остальные — её сектор (medii sector). Медиана связана с сектором радиуса поперечной радиомедиальной жилкой (radiomedial). Затем расположены кубитальные (cubitus), анальные (analis) и наконец югальные (jugum) жилки.
У стрекоз, подёнок, сетчатокрылых и немногих других, относительно примитивных, насекомых почти все промежутки между основными жилками крыла заняты густой сетью более мелких жилок и анастомозов (так называемая первичная сеть — архедиктион), а у прямокрылых и родственных форм особенно сильного развития достигает югальная область крыла. Способная собираться в мелкие складки, как гармошка, эта область мгновенно развёртывается роскошным веером. Обнажаемая на взлёте, нередко окрашенная в яркие цвета и как бы вспыхивающая в этот момент, она может отпугнуть преследователя неожиданностью эффекта.
У подавляющего большинства насекомых обычно развиты обе пары крыльев, хотя ведущая роль в полете, как правило, принадлежит передним. У некоторых насекомых, например у двукрылых, сохраняется лишь одна пара крыльев, а задние преобразуются в жужжальца — своеобразный «гироскоп автопилота», стабилизирующий полет. Обычно насекомые, сохранившие обе пары крыльев, функционально двукрылы: оба крыла с каждой стороны тела сцепляются в единую крыловую плоскость, и лишь стрекозы и немногие другие способны к автономным движениям каждого крыла.
Тенденция к функциональному объединению обеих пар крыльев или редукции одной из них обозначается как «принцип диптеризации», или «принцип Родендорфа—Шванвича», и проявляется почти в каждой группе высших насекомых. Ещё одно общее преобразование может быть названо «принципом костализации крыла», который выражается в смещении основных жилок вперёд, к косте, что соответствует последовательному совершенствованию лётных качеств. Несмотря на то что аэродинамические свойства крыльев насекомых исследованы весьма подробно и несмотря на притягательность идеи орнитоптера — летательного аппарата с машущими плоскостями, она всё ещё не реализована в должной мере в самолётостроении.
В практической работе систематика по характеру жилкования различают крылья с большим и малым числом жилок, а по их плотности — перепончатые, кожистые и роговые (рис. 31).
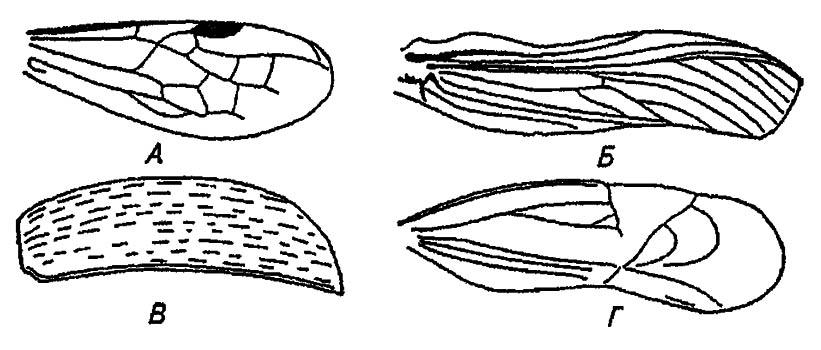
Рисунок 31. Различные типы крыла (по Бей-Биенко, 1966):
А — перепончатое; Б — кожистое; В — роговое; Г — полужёсткое надкрылье клопа
Сочленение крыла с телом представляет собой один из наиболее сложных и совершенных механизмов. Основания жилок крыла не доходят до края тергита, обеспечивая свободу движений крыловой пластинки. Здесь, в обширной сочленовной мембране, располагаются многочисленные склериты, ограничивающие и стабилизирующие возможные направления изгибаний. Некоторые склериты приводятся в движение мышцами и обеспечивают отведение и складывание крыла вдоль тела или изменяют его наклон. Лишь немногие насекомые держат крылья распростёртыми в стороны, что весьма ограничивает использование укромных убежищ и мест обитания.
Край тергита, от которого начинается сочленение крыла, имеет глубокую вырезку и две лопасти. Между ними помещаются 3–4 осевые, или аксиллярные, пластинки, с которыми, в свою очередь, связаны медиальные крыловые пластинки. Роль этих склеритов в складывании и расправлении крыльев можно понять из схемы, показанной на рисунке 32.
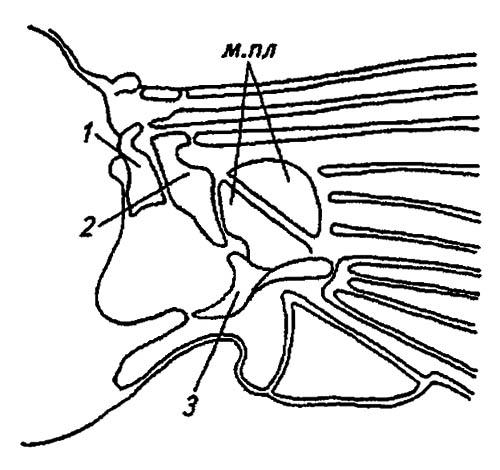
Рисунок 32. Сочленение крыла с грудью (по Шванвичу, 1949):
1, 2, 3 — первая, вторая и третья аксиллярные пластинки; м. пл — медиальные пластинки
Наиболее ранние исследования лётных качеств насекомых послужили основой для противопоставления планирующего полёта гребному и описания траектории движения крыльев. К. Марей (1869), прикрепляя к крылу кусочек полированного золота, наблюдал траекторию отражённого солнечного луча и описал наиболее характерные положения крыльев. С развитием техники эксперимента данные К. Марея были подтверждены; кроме того, было показано, что движения крыльев наряду с поддержанием тела в воздухе весьма своеобразно обеспечивают его продвижение вперёд. Если крыло закреплённого насекомого описывает замкнутую траекторию в виде отклонённой назад цифры 8, то при полете или в потоке воздуха внутри аэродинамической трубы траектория движения крыльев приобретает вид синусоиды, причём на её нижнем участке меняется наклон крыловой плоскости (рис. 33).
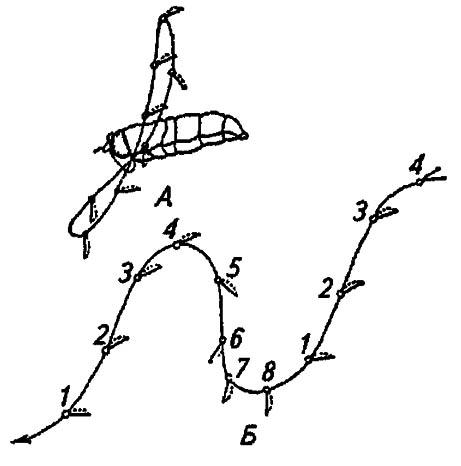
Рисунок 33. Траектория крыла насекомых (по Бей-Биенко, 1966):
А — при стоячем полете мухи-журчалки; В — при полёте вперёд; 1–8 — последовательные положения крыла
Опускаясь сверху вниз всей поверхностью, крыло противодействует падению тела. Поэтому данную часть траектории обозначают как элеваторную, а рабочей поверхностью крыла служит его нижняя плоскость. Далее, при подъёме крыла вверх, оно поворачивается вертикально, нижним краем вперёд, тем самым обеспечивая поступательное движение тела; рабочей поверхностью крыла при этом становится его верхняя сторона, а полет приобретает характер пропеллирующего. Чередование элеваторных и пропеллирующих ударов крыльями позволяет осуществлять весьма сложные маневры и в стремительном полёте вперёд, и при зависании на месте — способности, отличающей мух, бражников, пчёл и стрекоз.
У более примитивных насекомых элеваторная составляющая полёта явно преобладает: порхающие движения подёнок, веснянок и термитов не отличаются совершенством. В исходном состоянии полет, видимо, сводился к планированию или парашютированию — обширные крыловые плоскости древних насекомых, способные лишь к ограниченным движениям, содействовали их прыжкам и перелётам с ветви на ветвь деревьев.
Среди множества гипотез происхождения крыльев насекомых наибольшей популярностью пользуется «паранотальная теория». Торчащие в стороны боковые выросты тергитов грудных сегментов — паранотумы — могли со временем приобрести сочленения с телом и собственную мускулатуру. Сохранившиеся лишь у некоторых личинок и немногих взрослых форм, эти выросты отличали ныне вымерших насекомых, причём они формировались не только на средне- и заднегруди, но и на переднем грудном сегменте. До сих пор проблема происхождения крыльев и полёта насекомых не имеет однозначного толкования, порождая все новые и новые гипотезы.
Рассматривая челюсти грызущего ротового аппарата, мы убедились в их соответствии двуветвистым конечностям ракообразных и трилобитов. Однако в строении ног насекомых не остаётся никаких следов исходной двуветвистости. Они образованы следующими пятью члениками: тазиком (coxa), вертлугом (trochanter), бедром (femur), голенью (tibia) и лапкой (tarsus), подразделённой на более мелкие членики (рис. 34).
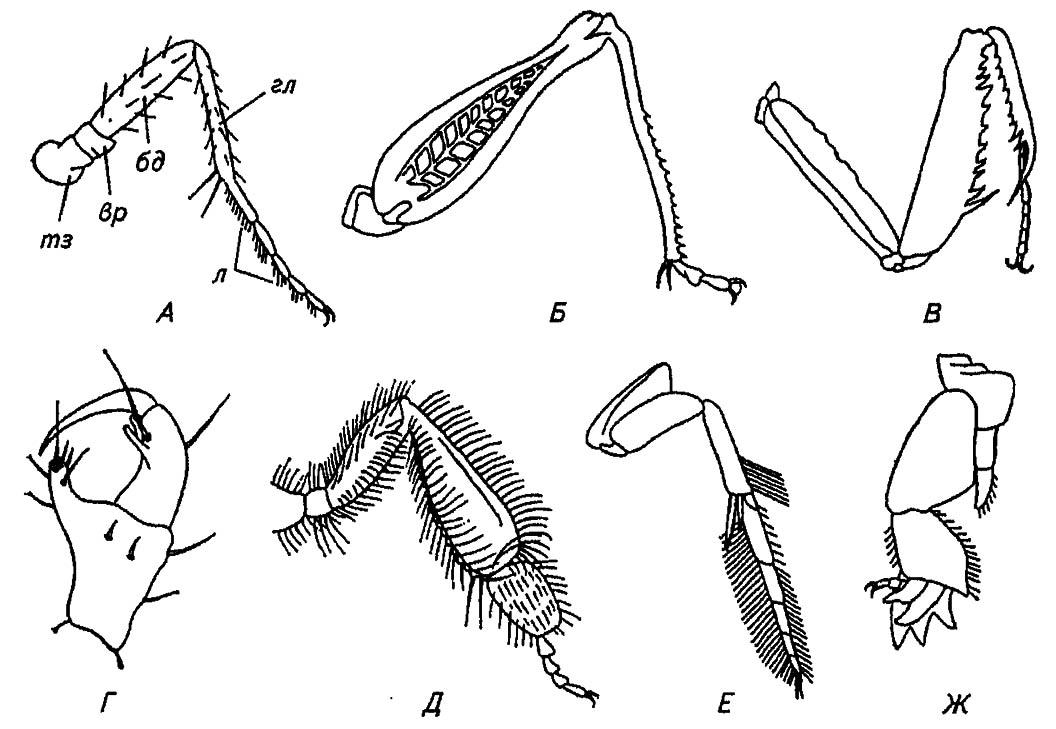
Рисунок 34. Различные типы ног насекомых (по разным авторам):
А — ходильная нога: тз — тазик; вр — вертлуг; бд — бедро; гл — голень; л — лапка; Б, В, Г, Д, Е, Ж — соответственно прыгательная, хватательная, прицепная, собирательная, плавательная, копательная нога
Тазик причленяется к плейриту посредством двух мыщелков, допускающих весьма ограниченные движения вперёд и вверх или назад и вниз (рис. 35).
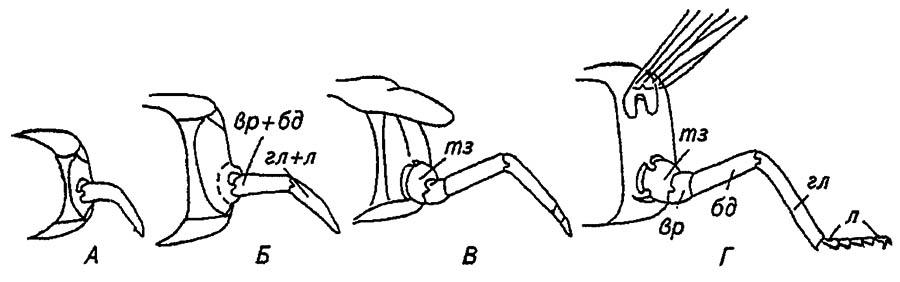
Рисунок 35. Формирование ноги насекомого в соответствии с субкоксальной теорией формирования плейрита (по Шванвичу, 1949)
А — ходильная нога: тз — тазик; вр — вертлуг; бд — бедро; гл — голень; л — лапка; Б, В, Г, Д, Е, Ж — соответственно прыгательная, хватательная, прицепная, собирательная, плавательная, копательная нога
Подвертлужное сочленение между бедром и вертлугом более подвижно. Обеспечивая движения вверх и вниз, оно наряду с предыдущим сочленением позволяет достичь любой точки поверхности, ограниченной лишь длиной конечности. Однако оба эти сочленения не позволяют приблизить конечность к телу. Для этого служит своеобразное коленное сочленение между бедром и голенью. Кроме того, имея горизонтальную ось вращения, оно не допускает перегиба на спинную сторону. В ещё большей степени ограничена подвижность вертлуга по отношению к бедру и лапки по отношению к голени. Впрочем, лапка, образованная несколькими (обычно пятью) члениками, выполняет особые функции. Заканчиваясь предлапкой (praetarsus) — разного рода коготками и присосками, — она предназначена для фиксации конечности на субстрате, что обеспечивает устойчивость и цепкость самого насекомого. Обычно на конце лапки имеется по два коготка с расположенными под ними присосками, или пульвиллами (pulvilli), и непарным эмподием (рис. 36).
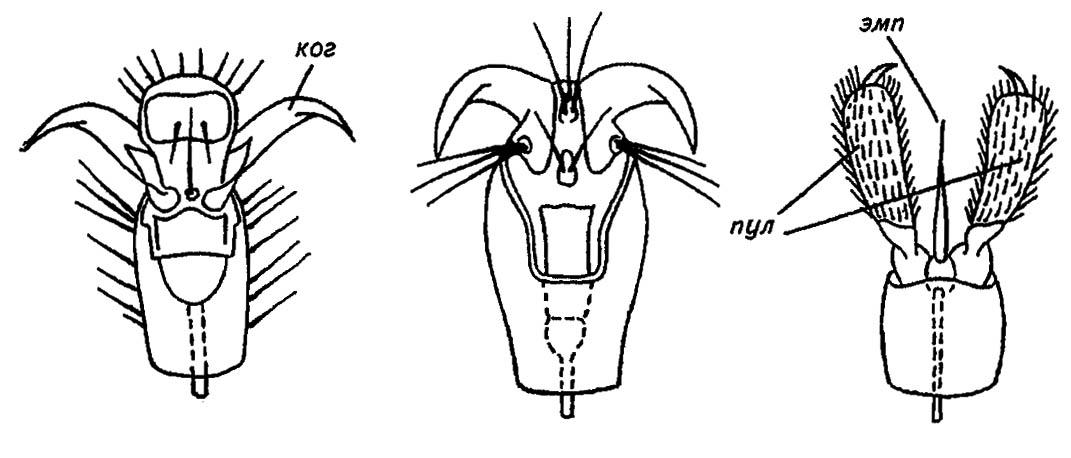
Рисунок 36. Концевой членик лапки (предлапки) у насекомых (по Шванвичу, 1949):
ког — коготки; пул — пульвиллы; эмп — эмподиум
Наглядно представить роль этих структур можно, наблюдая за движениями мухи по потолку или прилагая усилия, чтобы снять прицепившихся насекомых с растения или шерсти животного.
Особенно своеобразны прицепные подушечки кровососущего клопа Rhodniusprolixus St., образованные множеством волосков с косо срезанными вершинами (рис. 37).
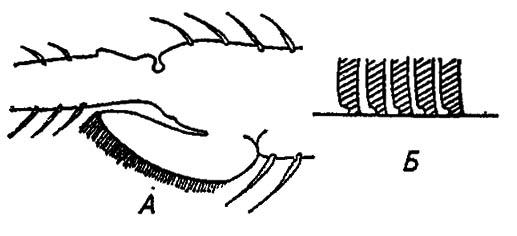
Рисунок 37. Присасывательные подушечки и волоски на голени клопа Rhodnius prolixus (по Шваивичу, 1949):
А — продольный срез через подушечку; Б — мениски между концами волосков и гладкой поверхностью субстрата
Благодаря силам поверхностного натяжения выступающих из волосков капелек клоп свободно ползёт вверх по зеркальному стеклу, но в положении вниз головой сразу утрачивает цепкость. В первом случае мениски капелек растягиваются и силы поверхностного натяжения возрастают; в обратном положении поверхностное натяжение сокращается и клоп соскальзывает вниз, как на коньках.
Разнообразию образа жизни насекомых и освоенных ими сред обитания соответствует разнообразие конечностей (см. рис. 34). Например, расширенные и уплощённые передние ноги жуков-навозников, личинок певчих цикад или медведок приспособлены к копанию и именуются копательными. При этом у роющихся в рыхлой земле медведок основание лапки — причленяется к основанию голени, имеющей выступающие зубцы. Эта конструкция функционирует наподобие сенокосилки или машинки для стрижки волос: тонкий корешок или стебель растения, ущемлённый между зубцами голени, легко срезается вращающейся лапкой. Мощное развитие задних ног у кузнечиков или саранчи соответствует способности к прыжкам, и поэтому их конечности называют прыгательными. Следует также упомянуть специализированные хватательные конечности богомолов, некоторых сетчатокрылых и клопов, плавательные ноги водных жуков и клопов, прицепные ноги паразитов, собирательные — пчёл и шмелей, прядильные — эмбий. Существуют и другие типы ног, особенности которых отражены в названиях. Однако наиболее характерны для насекомых обыкновенные ходильные или бегательные нош, свойственные мухам, тараканам, жужелицам и др. Конечности этого типа обеспечивают перемещение насекомых по суше, иногда на большие расстояния.

