Голова и её придатки
Голова насекомого (рис. 6) представляет собой плотную капсулу — эпикраниум, к которой причленяются ротовые придатки и антенны. Её поверхность разделена швами на участки и с обеих сторон имеет выпуклые фасеточные глаза. Номенклатура отдельных швов и участков заимствована из анатомии человека и часто используется для определения насекомых в практической работе систематика.

Рисунок 6. Схема строения головы насекомых спереди (А), сбоку (Б), сзади (В) и снизу (Г) (по Snodgrass, 1935):
гл — глаза; ант — антенны; глз — глазки; в. г — верхняя губа; мд — мандибулы; мк — максиллы; н. г — нижняя губа; т. ш — теменной шов; л. ш — лобные швы; л — лоб; нал — наличник; щ — щёки; пщ — подщёки; зщ — защёки; зт — затылок; ззт — заднезатыпок; з. о — затылочное отверстие; р. от — ротовое отверстие; т. я — тенториальные ямки; гпф — гипофаринкс; з. ш — затылочный шов; ззт. ш — заднезатылочный шов; ш.скл — шейные склериты; сл. п — отверстие слюнного протока
Обычно самая верхняя часть эпикраниума представлена теменем (Vertex), разделённым срединным эпикраниальным швом на правую и левую половины. Спускаясь на переднюю лицевую поверхность головы, эпикраниальный шов раздваивается налобные швы, ограничивающие треугольный лоб (frons).
По обеим сторонам лба располагаются щёки (genae), переходящие без видимых границ в темя. Сзади они ограничиваются затылочным швом, служащим также задней границей темени. Позади затылочного шва за щеками имеются узкие защёки (postgenae), а за теменем — поле затылка (occiput). В свою очередь, и затылок, и защёки отделены заднезатылочным, или постмаксиллярным, швом от заднезатылка (postocciput), окаймляющего узким воротничком затылочное отверстие. На лицевой поверхности и с боков головной капсулы нижней границей лба, щёк и защёк является шов, именуемый подолбом эпистомальным швом, под щеками субгенальным и под защёками гипостомальным швом. Он отделяет ото лба широкий наличник (clypeus), с которым сочленена верхняя губа (labrum), а от щёк и защёк узкие подщёки (subgenae) с верхними (mandibulae) и нижними (maxillae) челюстями насекомого. Непосредственно под заднезатылком причленяется нижняя губа (labium).
На лбу и темени многих насекомых находятся 1–3 маленьких дорзальных, или теменных, глазка, а у концов эпистомального и постмаксиштярного швов, то есть между лбом и наличником и над основаниями нижних челюстей, — глубокие тенториальные ямки. Эти ямки образованы втачиваниями покровов, которые проникают узкими рукавами внутрь головной капсулы и формируют её внутренний скелет — тенторий (tentorium), показанный на рисунке 7.
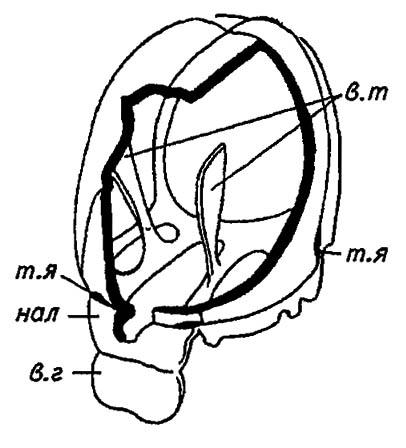
Рисунок 7. Внутренний скелет головы (по Шванвичу, 1949):
нал — наличник; в. г — верхняя губа; т. я — тенториальные ямки; в. т — ветви тентория
И тенторий, и швы эпикраниума придают голове насекомых особую прочность и служат опорой для мощных челюстных мышц и мышц глотки. Этим же целям отвечают общая консолидация и слияние головных сегментов друг с другом, не допускающие взаимных смещений и деформаций.
Основываясь на наиболее очевидных признаках сегментации, а именно на наличии придатков и межсегментных борозд, можно выделить в голове насекомого сегменты нижней губы (лабиальный, или постмаксиллярный, сегмент) и нижних челюстей (максиллярный сегмент), ограниченные заднезатылочным и затылочным швами эпикраниума. Далее выделяется сегмент верхних челюстей (мандибулярный сегмент), который, однако, не отделён швом от впередилежащего, премандибулярного, сегмента. Лишённый развитых придатков и собственных границ, этот сегмент выделяется на основании парных целомов и зачатков придатков, формирующихся у эмбриона, но редуцирующихся впоследствии (рис. 8).
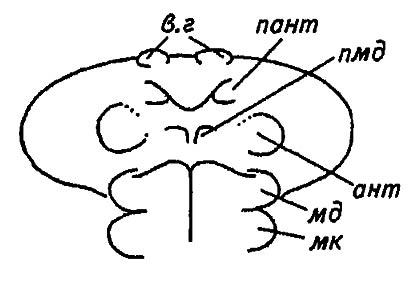
Рисунок 8. Формирующиеся сегменты головы зародыша палочника:
в. г, пант, ант, пмд, мд, мк — зачатки сегментов верхней губы, преантенн, антенн, премандибулярного, мандибулярного и максиллярного сегментов соответственно
Возможно, что эти придатки соответствуют вторым антеннам ракообразных. Далее следуют сегменты антенн и преантенн, причём придатками последнего является верхняя губа, образованная слившимися друг с другом парными зачатками. Считая, что акрону членистоногих и предротовой лопасти кольчатых червей соответствует фронто-клипеальная область головы насекомого (область лба и наличника), мы, таким образом, насчитываем шесть сегментов головного отдела, признаки которых выражены в разной степени.
Приведённая трактовка сегментарного состава головы насекомых не является общепринятой. Некоторые авторы насчитывают в голове насекомых до семи отдельных сегментов, другие — сокращают их число до пяти и даже четырёх. До сих пор нет единой точки зрения о производных акрона и о придатках преантеннального сегмента, о природе его целомов и соответствии антенн либо туловищным конечностям, либо придаткам акрона. Эти вопросы имеют принципиальное значение для решения проблемы происхождения насекомых. Разнообразие структур головной капсулы насекомых, её форм и пропорций не поддаётся сколько-нибудь схематичному и компактному описанию. Поэтому мы ограничимся рассмотрением лишь немногих характерных примеров, приведённых на рисунке 9.
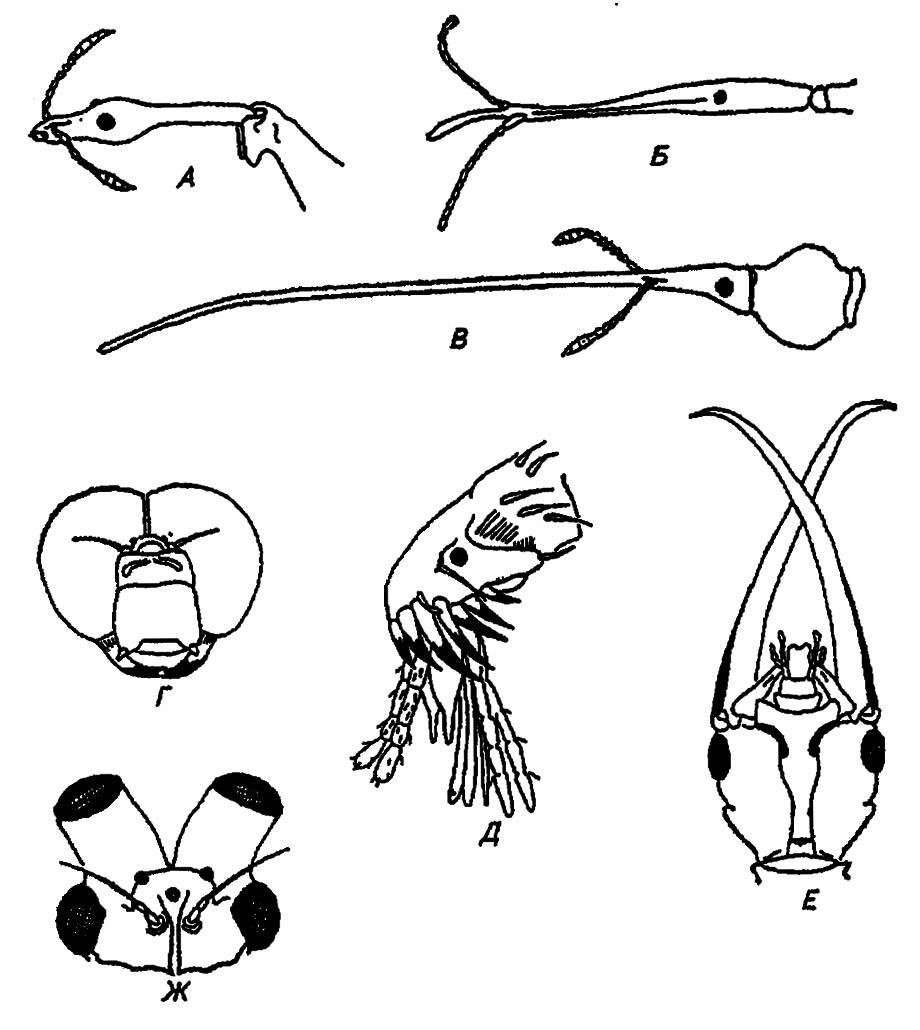
Рисунок 9. Отдельные примеры видоизменения головы у насекомых:
А — жук Apoderus longiceps; Б — жук Brenthis sp.; В — жук Anthiarrhinus rectirostis; Г — стрекоза; Д — блоха; Е — личинка коридала (Neuroptera); Ж — подёнка (по разным авторам)
Особенно сильные изменения ротовых органов, глаз и антенн. На некоторых из них мы остановимся впоследствии, сейчас укажем только на различные способы постановки головы и её сочленения с туловищем. Наиболее типичное состояние — гипогнатическое, при котором продольная ось эпикраниума перпендикулярна оси тела, а ротовые органы обращены вниз.
При более специализированном прогнатическом типе, свойственном многим хищникам, ротовые органы обращены вперёд и ось эпикраниума совпадает с осью тела. Наконец, при опистогнатическом типе (у цикад и некоторых других насекомых) голова сильно скошена вниз и назад, а её ось образует острый угол с осью тела (рис. 10).
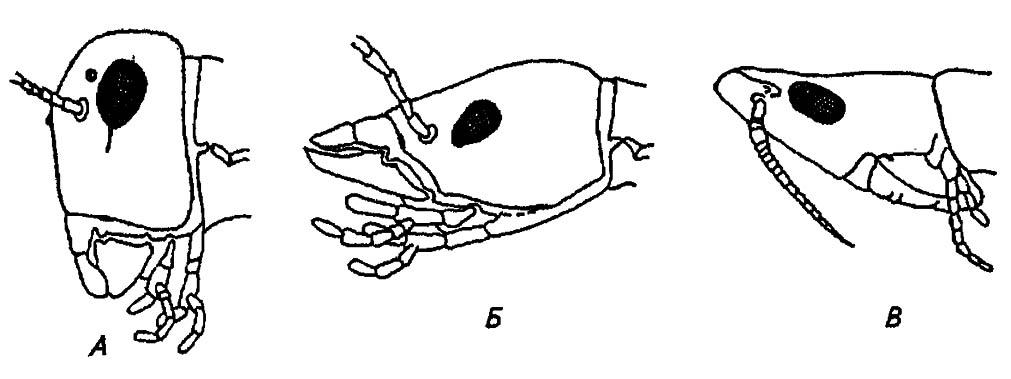
Рисунок 10. Различные типы постановки головы у насекомых (по Шванвичу, 1949):
А — гипогнатаческий; Б — прогнатический; В — опистогнатический
Антенны насекомых представляют собой парные членистые придатки, обычно весьма подвижные и хорошо развитые. Лишь у личинок мух и некоторых перепончатокрылых, а также у самок веерокрылых и бабочек-мешочниц антенны редуцированы до небольших бугорков или дисков. Выполняя роль своеобразных локаторов, они могут считаться по совокупности отдельных рецепторов органами осязания и обоняния, а иногда и органами слуха, воспринимающими ультразвук. Полагают, что они соответствуют первой паре антенн ракообразных.
Положение антенн, как и их размеры и форма, весьма изменчиво у разных видов. Обычно антенны располагаются в неглубокой антеннальной ямке на темени, вблизи глаз или верхних челюстей насекомого и состоят (рис. 11) из основного членика (scapus), членика-ножки (pedicellum) и многочленикового жгута (flagellum).
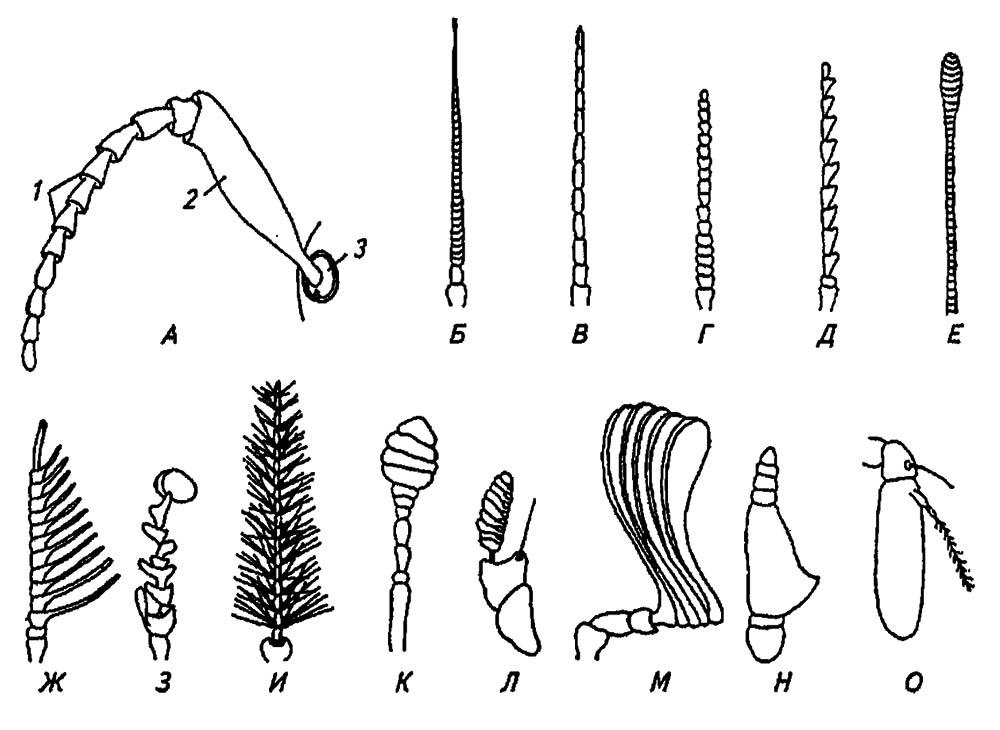
Рисунок 11. Антенны насекомых:
А — общий план строения: 1 — жгут; 2 — ножка; 3 — основной членик. Антенны: щетинко-видная (Б); нитевидная (В); чётковидная (Г); пиловидная (Д); булавовидная (Е); гребенчатая (Ж); неправильная (З); перистая (И); головчатые жуков-мертвоедов (К), блох (Л); пластинчатая (М); с расчленённым члеником (у двукрылых) (Н) и щетинконосная (О) (по разным авторам)
Подвижность основного членика обеспечивается мышцами, связанными с тенторием, однако он имеет и собственную мускулатуру, управляющую движениями ножки и жгута, членики которого обычно лишены мышц. Лишь у некоторых, наиболее примитивных насекомых членики жгута снабжены собственными мускулами и способны к самостоятельным движениям.
Нередко строение антенн используется в определении насекомых, и поэтому в практической систематике существуют специальные обозначения для разных их типов (см. рис. 11). Наиболее просты нитевидные и щетинковидные антенны, различающиеся тем, что последние утончаются к вершине. Далее выделяют чётковидные антенны, жгут которых напоминает связку бус, пиловидные (одно- и двусторонние), гребенчатые, булавовидные, веретеновидные, пластинчатые и перистые. Наряду с ними различают коленчатые антенны с весьма длинным основным члеником, к которому под углом прикрепляются ножка и жгут, а также щетинконосные и неправильные. У некоторых видов насекомых антенны у самцов развиты сильнее, чем у самок, что связывают с необходимостью активного поиска последних, ведущих менее подвижный образ жизни. Подчас антенны принимают на себя, казалось бы, несвойственные им функции, например обеспечение дыхания и удержание добычи (жуки и личинки водолюбов) ил и органов равновесия (клопы-гладыши).
Характеризуясь широчайшим разнообразием вариантов, ротовые органы насекомых в своей основе представлены грызущим ротовым аппаратом, свойственным прямокрылым (Orthoptera) и многим другим, относительно примитивным представителям класса (см. рис. 6). Напомним, что они являются придатками сегментов головы и, исключая непарную верхнюю губу, гомологичны парным туловищным конечностям. Верхняя губа, хотя и развивается из парных зачатков, считается либо новообразованием, либо преобразованными в ходе эволюции придатками акрона. Обычно она имеет вид широкой лопасти, сочленённой с наличником и несущей на своей внутренней поверхности своеобразные зубцы эпифаринкса, или надглоточника (рис. 12).
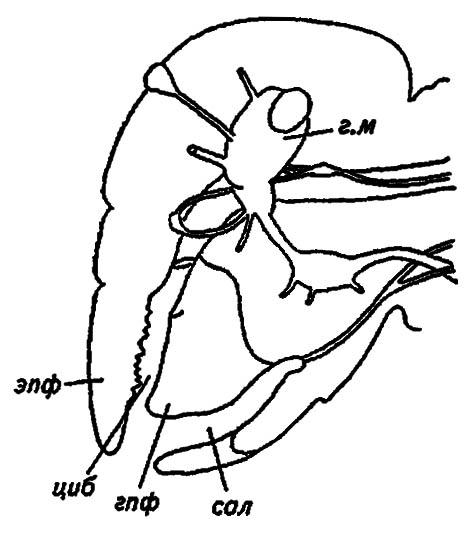
Рисунок 12. Голова насекомого в разрезе (по Шванвичу, 1949):
г. м — головной мозг; эпф — эпифаринкс; гпф — гипофаринкс; циб — цибарий; сал — саливарий
Верхние челюсти (mandibulae), или жвалы, показанные на рисунке 13, А, — придатки мандибулярного сегмента, играют основную роль в грызущем ротовом аппарате, откусывая и сминая частички пищи. В соответствии с этим у хищных насекомых они особенно прочны и имеют форму серпа, узкого и острого у вершины, но массивного и тупого у основания. Сочленяясь с головной капсулой двумя мыщелками, они движутся в одной плоскости. У растительноядных форм жвалы более тупые и зазубренные и снабжены в основной своей части (коренной, или молярной) серией рёбер и борозд, содействующих перетиранию пищи. У примитивных насекомых жвалы имеют лишь одно сочленение и способны к менее точным и сильным движениям в разных плоскостях. Они непохожи на ноги насекомого, хотя и являются их гомологами, как парные придатки сегмента. Нижние челюсти (maxillae) грызущего ротового аппарата наиболее сложны по строению (рис. 13, В). Они образованы основным члеником (cardo), стволиком (stipes), парой жевательных лопастей — наружной (galea) и внутренней (lacinia) — и членистым нижнечелюстным щупиком (palpus maxillaris), отходящим от стволика. Приводимые в движение системой мышц, все эти компоненты нижней челюсти принимают участие в пережёвывании пищи, причём нижнечелюстной щупик является и органом вкуса. При сравнении его с двуветвистыми конечностями трилобитов и ракообразных можно прийти к мнению, что нижнечелюстной щупик отвечает их внутренним ветвям и, по-видимому, гомологичен придаткам грудных сегментов насекомого, то есть ногам. Вместе с тем есть основания сравнивать жевательную лопасть — лацинию — с мандибулой, так как жвалы некоторых многоножек имеют развитые стволик и основание, а у ряда ракообразных они сохраняют верхнечелюстной щупик.
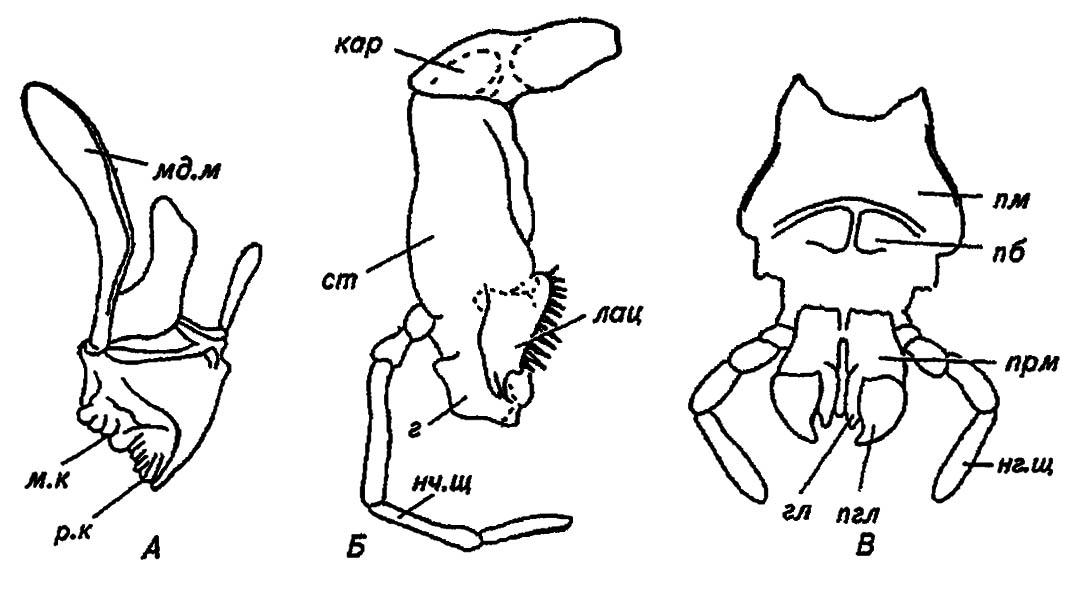
Рисунок 13. Ротовой аппарат грызущего типа (по Gillot, 1980):
А — верхние челюсти (мандибулы): мд. м — мандибулярные мыщелки; м. к — молярный (коренной) край; р. к — режущий край мандибулы; Б — нижние челюсти (максиллы): кар — кардо (основной членик); ст — стипес (стволик); г — галеа; лац — лациния; нч. щ — нижнечелюстной щупик; В — нижняя губа: пб — подбородок (ментум); пм — постментум; прм — прементум; гл — глоссы; пгл — параглоссы; нг. щ — нижнегубной щупик
Нижняя губа (labium), по существу, является второй парой нижних челюстей, но сросшихся друг с другом в основании. Действительно, каждому компоненту нижней челюсти соответствует определённая структура в нижней губе (рис. 13, В). Например, основным членикам максилл отвечает постментум (postmentum), разделённый на гомологичный стволикам подбородок (mentum) и подподбородок (submentum). Сопоставление жевательных лопастей с придатками прементума (praementum) позволяет соотнести лацинии с язычками (glossae) нижней губы, а галеа — с придаточными язычками (paraglossae). Сходство нижнегубных щупиков (palpi labiales) с нижнечелюстными очевидно и не требует комментариев.
Итак, грызущий ротовой аппарат образован верхней губой, двумя парами челюстей и нижней губой. Расположенные вокруг рта, они ограничивают предротовую полость, в которую в виде толстого и мягкого языка вдаётся подглоточник (hypopharynx) (см. рис. 12). Разделяя предротовую полость на два отдела — передний и задний, подглоточник противополагается надглоточнику (epipharynx) и вместе с ним участвует в сминании пищевого комка перед заглатыванием. В передний отдел, называемый цибарием (cibarium), открывается глотка, в задний, или саливарий (salivarium), — протоки слюнных желез.
Такова принципиальная схема грызущего ротового аппарата, с помощью которого насекомые, им обладающие, питаются твёрдой пищей, откусывая и дробя её мощными челюстями. Вместе с тем грызущий ротовой аппарат служит общей основой для всех возможных модификаций ротовых органов насекомых. В конечном счёте и хоботки бабочек-бражников, и острые стилеты клопов, и сверла мухи цеце и жигалок основаны на примитивных ротовых органах грызущего типа, свойственных и тараканам, и термитам, и прямокрылым, и веснянкам, и многим другим относительно примитивным насекомым.

