Покровы тела и их производные
Покровы насекомых обладают различными свойствами, которые позволяют им наряду с опорной (механической) функцией наружного скелета выполнять и другие сложные функции. Особенно важны их барьерные свойства, регулирующие взаимодействие организма с внешним миром и обеспечивающие постоянство его собственной внутренней среды. Покровы служат также носителями окраски тела, местом отложения резервных и некоторых конечных продуктов метаболизма, средоточием рецепторов и желез.
Основу покровов составляет гиподерма — однослойный эпителий, подостланный изнутри базальной мембраной и покрытый снаружи кутикулой (рис. 56). Бесструктурная базальная мембрана секретируется гемоцитами — клетками «крови» насекомых и состоит главным образом из мукополисахаридов. Клетки гиподермы одноядерны, имеют на поверхности многочисленные микроворсинки и нередко преобразуются либо в эноциты — крупные секретирующие клетки, либо в компоненты гиподермальных желез, либо в сенсиллы — разнообразные волоски и щетинки, столь характерные для внешнего облика насекомых.

Рисунок 56. Покровы тела насекомых (по Gillot, 1980; Шванвичу, 1949):
гип — клетки гиподермы; эндк — эндокутикула; экзк — экзокутикула; эпк — эпикутикула; ч. вол — чувствительный волосок; баз. м — базальная мембрана; жел. кл — железистая клетка
Выделяемая гиподермой кутикула — относительно косное, не имеющее собственного метаболизма образование, пронизанное многочисленными поровыми каналами. Эти каналы, отходящие от клеток, обеспечивают транспорт веществ при секреции отдельных слоев кутикулы и при регенерации покровов.
Обычно кутикула состоит из двух слоев: прокутикулы и эпикутикулы, которые обеспечивают её прочность и эластичность, а также химическую стойкость и непроницаемость. У водных и почвообитающих форм эпикутикула развита слабо или отсутствует, но у обитателей наземной среды она отличается сложнейшей дифференциацией. Прокутикула в химическом отношении представлена связанным с белками хитином — высокомолекулярным азотсодержащим полисахаридом (поли-N-ацетил-?-глюкозамин), близким по структуре к гликогену и целлюлозе (рис. 57).
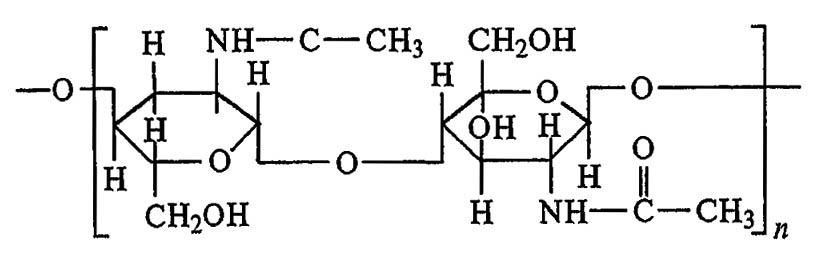
Рисунок 57. Структурная формула поли-N-ацетил-α-глюкозамина
Нерастворимый ни в воде, ни в щелочах, ни в органических растворителях хитин разлагается в крепких неорганических кислотах и при длительном нагревании с едким кали частично преобразуется в хитозан. Последний в присутствии слабых растворов серной кислоты даёт фиолетовое окрашивание с йодом, что используется как качественная реакция на хитин. В организме насекомого хитин синтезируется, по-видимому, из глюкозы и разлагается в природе лишь немногочисленными видами бактерий и грибов, выделяющих фермент хитиназу.
Водорастворимые фракции белков кутикулы, названные артроподином, образуют вместе с хитином основной глюко-протеиновый комплекс. Из нерастворимых в воде фракций наиболее своеобразен белок — резилин, накапливающийся в наиболее эластичных участках покровов, например в местах сочленения крыльев с телом. Наряду с хитином, составляющим до половины сухой массы кутикулы, и белками в её состав входят липиды, производные фенола, ферменты и небольшое количество минеральных соединений.
Прокутикула насекомых обычно подразделяется на экзо- и эндокутикулу. Последняя эластична и прозрачна. Она образована тончайшими слоями, перекрывающиеся фибриллы которых придают ей прочность и гибкость. Внутри эндокутикулы могут быть полости, рассечённые опорными балками, перегородками и колонками. Экзокутикула уступает по толщине эндокутикуле и особенно прочна. В местах сочленений она сильно утончена или совсем редуцируется. Особые свойства экзокутикулы определяются переходом ее водорастворимых белков (артроподинов) в нерастворимые склеротины. Такого рода склеротизация, сравнимая с технологическим процессом дубления кож, осуществляется при участии производных фенола и регулируется специальным гормоном — бурсиконом, выделяющимся в последних ганглиях брюшной нервной цепочки.
У некоторых насекомых имеется также переходный слой — мезокутикула с меньшим содержанием склеротинов, и слой Шмидта — узкая полоска аморфной, свежевыделенной и ещё не стабилизированной эндокутикулы (рис. 58).

Рисунок 58. Генезис кутикулы. Последовательные фазы (7–6) аполизиса и формирования новой кутикулы (по Gillot, 1980):
прк — прокутикула; Шм — слой Шмита; эноц — эноциты; пор. к — поровый канал; экзв. ж — экзувиальная жидкость; эли, экз, энд — эпи-, экзо-, эндокутикула
В сформированном состоянии кутикула не только прочна, но и мало проницаема для воды. Это её свойство приобретает особое значение для сухопутных насекомых, испытывающих из-за малых размеров тела постоянную угрозу иссушения: ведь отношение площади поверхности тела к его объёму у них иное, нежели у крупных животных, и площадь испарения воды относительно большая. При этом основную роль в поддержании водного баланса играет тончайшая эпикутикула. Продуцируемая отчасти клетками гиподермы, отчасти специализированными железами, она, в свою очередь, подразделяется на четыре слоя: кутикулиновый, полифеноловый, восковой и цементный.
Внешний цементный слой, развитый не у всех насекомых, по составу близок к шеллаку (лаккозо-липоидной смеси). В следующем за ним, восковом, слое преобладают эфиры жирных кислот. В нем можно выделить несколько зон. Самый внутренний мономолекулярный слой состоит из липидов, ориентированных гидрофобными (неполярными) группами наружу, что придаёт его поверхности высокий контактный угол с молекулами воды и устойчивость к иссушению. В среднем, менее упорядоченном слое находится большая часть воска, пропитанного веществами цементного слоя. Наконец, внешняя зона воскового слоя образует характерные для некоторых насекомых восковые налёты и структуры.
Полифеноловый слой, по-видимому, не имеет значения самостоятельной структуры и представляет собой задубленную часть кутикулинового, прилегающего к экзокутикуле слоя. Последний развит по всей поверхности тела, заходя также в трахеи, во впячивания покровов и в эктодермальные производные кишечника и полового тракта. Лишь на отдельных участках сенсилл и желез кутикулиновый слой не развит. Представляя собой основу эпикутикулы, он выполняет роль селективного барьера проницаемости и «активирующего фактора» для ферментативного гидролиза эндокутикулы при линьке. Вместе с тем кутикулиновый слой проницаем для воска, а в некоторых случаях и для воды. Подстилая восковой «монослой», он определяет ориентацию его молекул и значение контактного угла, текстуру поверхности кутикулы. Кроме того, будучи весьма жёстким, он ограничивает растяжение покровов в целом. Однако, несмотря на многообразие и важность исполняемых функций, о его составе известно только то, что кутикулин включает липопротеиды, задубливающиеся в полифеноловом слое, и, по-видимому, секретируется при участии эноцитов.
Для большинства наземных насекомых скорость испарения воды из тела не превышает 1% её общих запасов [1–3 мл/(см2 х ч)], при этом испарение происходит в основном из трахейной системы и в гораздо меньшей степени через покровы. Однако непроницаемость покровов неабсолютна, и некоторые химические соединения с большей или меньшей скоростью достигают внутренней среды насекомого за счёт либо пассивного, либо активного переноса их молекул через кутикулярный барьер.
Пассивный перенос молекул не связан с биологической активностью гиподермы и определяется лишь законами диффузии и осмоса, концентрацией проникающих растворов, сорбционными свойствами и электрическим зарядом поверхности. При активном переносе, осуществляемом иногда вопреки градиенту концентрации веществ, в направлении, противоположном действию осмотических сил, необходимая для транспорта энергия высвобождается в обменных процессах клеток гиподермы. Проблема переноса молекул сквозь кутикулу имеет важное значение при разработке химических средств борьбы с вредителями, поскольку не все пестициды способны проникать в тело насекомого через его покровы.
Экспериментально было показано, что проницаемость кутикулы сохраняется лишь до некоторой критической точки, до температуры, соответствующей началу плавления восков. При удалении воскового слоя растворителями или абразивными материалами проницаемость сразу возрастает и насекомое теряет способность противостоять иссушению. На основе этого можно объяснить «эффект Дилера» — гибель амбарных вредителей при опудривании охраняемых от них запасов зерна неядовитыми, но нарушающими целостность воскового слоя эпикутикулы порошками.
Однако кутикулярные воска ограничивают проницаемость покровов насекомых только в одном направлении, от прокутикулы наружу. Транспорт веществ внутрь тела происходит быстрее и легче, и именно благодаря данной функциональной асимметрии кутикулы становится возможным активное поглощение воды из влажной, но не насыщенной водой атмосферы.
По мнению Д. Бимента, исходная проницаемость кутикулы обусловлена упорядоченностью воскового мономолекулярного слоя, молекулы которого, располагаясь на задубленном кутикулине под углом около 25°, обращены своими полярными (гидрофильными) группами внутрь, а неполярными (гидрофобными) — наружу. Удерживаемые в этом положении силами Ван-дер-Ваальса, они сообщают гидрофобность всей поверхности тела. Если молекулы ориентируются иначе, то кутикула становится более гидрофильной и пропускает воду. При увеличении температуры возрастает кинетическая энергия молекул, которые, теряя связи друг с другом, открывают пространства для проникновения молекул воды.
Собственно процесс переноса молекул воды через покровы насекомых, по мнению Д. Бимента, осуществляется белками. Поглощая из растворов воду, в соответствии с их способностью к гидратации при данной изоэлектрической точке они стремятся к достижению равновесного состояния. Под воздействием внешних источников энергии состояние, а следовательно, и степень гидратации белковых молекул изменяются, что может привести к поглощению ими новых порций воды. Выключение источника энергии сразу восстанавливает равновесное состояние молекул, и они отдают поглощённую воду. Насасывая при гидратации воду с обеих сторон, белки прокутикулы при дегидратации отдают её внутрь тела. Повторение циклов гидратация — дегидратация при периодическом воздействии позволяет реализовать водообеспечение организма.
Разные соединения проникают через кутикулярные барьеры с различной скоростью, а некоторые вообще не проникают через них. Иногда при совместном нанесении на покровы смесей плохо проникающих веществ может возникнуть неожиданный эффект резкого повышения проницаемости кутикулы. В токсикологии данное явление рассматривается как пример «потенцированного синергизма ядов» и используется для повышения эффективности инсектицидов. Однако конкретные механизмы проникновения инсектицидов сквозь покровы изучены пока недостаточно.
Все контактные пестициды условно разделяют на две группы: соединения, не нарушающие кутикулу, и соединения, разрушающие те или иные её структуры и меняющие проницаемость. К первой группе относят многие фосфорорганические инсектициды, ко второй — хлорорганические, но следует помнить об искусственности данного противопоставления и о специфичности покровов насекомых, допускающих применение избирательно действующих средств отравления.
Вместе с тем имеющиеся у насекомых дыхальца трахейной системы, проницаемые пористые рецепторные мембраны, функциональная асимметрия кутикулы могут существенно облегчить проникновение инсектицидов в гемолимфу, полость тела, нервную систему насекомых. Кроме того, все насекомые по мере развития претерпевают линьки и, сбросив старую шкурку, становятся уязвимы для разного рода воздействий.
Незадолго до наступления линьки клетки гиподермы увеличиваются, приобретают заметные ядрышки, многочисленные рибосомы и, подготовившись таким образом к интенсивному синтезу и секреции, отслаивают от себя кутикулу. В этот момент, получивший название аполизиса, между клетками гиподермы и кутикулой проникает неактивный линочный гель. Одновременно клетки гиподермы выделяют тончайший слой кутикулина, появление которого служит сигналом к началу растворения старой эвдокутикулы. Содержащая протеиназы и хитиназу линочная жидкость не действует на склеротины экзокути-кулы и защищенные кугикулином клетки гиподермы, но интенсивно разлагает эндокутикулу.
Растворяясь как кусок подтаивающего льда, старая кутикула становится все более тонкой, но в то же время начинается выделение новой кутикулы. При секреции кутикул и на особой активностью отличаются эноциты, которые, видимо, синтезируют его липопротеидовых предшественников. Образующийся кутикулиновый слой, первоначально ровный и гладкий, вскоре приобретает морщинки и складки, что обеспечивает возможность последующего увеличения размеров тела. Лишь после этого вслед за секрецией полифенолов происходит задубливание внешнего слоя кутикулина и начинается выделение новой прокутикулы.
Наращивая новые слои, прокутикула становится более толстой, а отходящие от клеток гиподермы тонкие цитоплазматические тяжи дают начало поровым каналам. По мере их формирования начинается отложение воскового слоя эпикутикулы. Секретируемые клетками гиподермы как водно-жировые жидкие кристаллы его компоненты проходят по поровым каналам на поверхность; у некоторых насекомых этот процесс длится всю жизнь. Наконец, наступает экдизис. Этим термином в отличие от аполизиса отмечают момент сбрасывания старой кутикулы, от которой в виде линочной шкурки, или экзувия, сохраняются только экзокутикула и эпикутикула (см. рис. 58). В момент разрыва экзувия, обусловленного заглатыванием воздуха (или воды у водных форм) и сильным сокращением мышц брюшка, создающим повышенное давление в передней части тела, линяющее насекомое расправляет новую мягкую кутикулу и придатки.
Рисунок 58. Генезис кутикулы. Последовательные фазы (7–6) аполизиса и формирования новой кутикулы (по Gillot, 1980):
прк — прокутикула; Шм — слой Шмита; эноц — эноциты; пор. к — поровый канал; экзв. ж — экзувиальная жидкость; эли, экз, энд — эпи-, экзо-, эндокутикула
Продолжающаяся секреция воска и формирование новых слоев эндокутикулы и цементного слоя сопутствуют указанным явлениям, однако наиболее важное событие после экдизиса — это склеротизация покровов.
На экспериментальных моделях было показано, что, в частности, при формировании пупария высших мух этот процесс состоит из следующих этапов. Накапливающийся в гемоцитах тирозин окисляется до дигидроксифенилаланина (сокращённо — ДОФА), который после декарбоксилирования преобразуется в дофамин. Последний сосредоточивается в клетках гиподермы и здесь же превращается в N-ацетил-дофамин, который выходит на поверхность кутикулы по поровым каналам и под действием дифенолоксидазы окисляется до хинона. Диффундируя в прокутикулу, хиноны связывают молекулы белка и содействуют формированию склеротинов. Параллельно склеротизации покровов происходит изменение их окраски благодаря образованию меланинов. Эти процессы (склеротизация и меланизация покровов) развёртываются независимо друг от друга, но сопряжены во времени и в обоих принимают участие тирозин и продукты окисления ДОФА. Это можно проиллюстрировать упрощённой схемой, показанной на рисунке 59.

Рисунок 59. Схема химических превращений при склеротизации кутикулы:
I, II — протеин I и протеин II (по Gillot, 1980)
При затвердевании кутикулы меняются её физические свойства и структура. Сокращение гидрофильных групп, включающихся в формирование склеротинов, приводит к более компактному расположению компонентов прокутикулы, которая соответственно обезвоживается и становится более тонкой.
Все эти процессы регулируются специфическими гормонами. В частности, активируя тирозиназу в гемоцитах и увеличивая проницаемость клеточных мембран для тирозина и дофамина, уже известный нам бурсикон служит регулятором склеротизации и пигментации покровов. Экдизис и отложение слоев эндокутикулы и воска определяются гормонами головного мозга, а аполизис — линочным гормоном, продуцируемым в переднегруди.
Рассмотрим структуры, в образовании которых принимают участие покровы тела. Важнейшие из них — это уже указанные выше элементы внутреннего скелета (аподемы, фрагмы, тенторий и др.), а также разнообразные выросты, щешнки и гиподермальные железы.

Рисунок 60. Кутикулярные выросты переднеспннки у цикадок-горбаток (А, Б, В) и жука-геркулеса (Г) (по Romoser, 1981)
Особенно причудливы листовидные или ветвящиеся выросты переднеспинки некоторых экзотических цикад и жуков, своеобразные рога и гребни (рис. 60). Более обычны волоски и чешуйки, характер распределения и форму которых нередко используют в систематике. Как специализированные органы чувств (сенсиллы) они формируются из двух клеток гиподермы (рис. 61). Одна из них — трихогенная клетка — выпускает сквозь пору в эндокутикуле цитоплазматический тяж и образует тело самой щетинки, другая — мембранная клетка — выделяет гибкую сочленовную мембрану вокруг основания щетинки.

Рисунок 61. Формирование крыловой чешуйки у бабочек (по Weber, 1966):
трих. кл — трихогенная клетка; мем. кл — мембранная клетка; гип — клетки гиподермы
Очень крупные и подвижные щетинки, расположенные на ногах, участвуют в движении и именуются шпорами. Однако наибольшей сложности строения достигают крыловые чешуйки бабочек, развивающиеся также из трихогенной и мембранной клеток гиподермы. В сформированном состоянии они представлены тончайшими решетчатыми и ребристыми пластинками (рис. 62), на которых происходят интерференция и дифракция световых волн и возникают яркие цветовые эффекты.

Рисунок 62. Тонкое строение крыловой чешуйки бабочки (по Romoser, 1981; Шванвичу, 1949)
Переходя к описанию гиподермальных желез, отметим, что все клетки гиподермы в определённые периоды жизни насекомого выполняют роль желез, секретирующих кутикулу. Здесь мы рассмотрим лишь специализированные железы: пахучие и ароматические железы клопов и пчёл; отпугивающие железы жуков-бомбардиров и термитов-солдат; ядовитые железы гусениц бабочек; восковые и лаковые железы равнокрылых; шёлкоотделительные железы эмбий; аллотрофические железы, предназначенные для вскармливания иных особей своего или чужого вида; железы, продуцирующие феромоны.
Перечисленные названия свидетельствуют о назначении желез или производимых ими секретов, а сами железы можно классифицировать как одноклеточные или многоклеточные, более или менее сложные, мелкие или заполняющие почти всю полость тела. Вместе с тем все железы имеют общие признаки строения: в основе их лежат секреторные элементы; перед их выделением наружу через различные протоки или системы протоков секреты накапливаются в резервуаре.

Рисунок 63. Пахучая железа нимфы клопа-соддатика (по Шванвичу, 1949):
1 — железистые клетки; 2 — полость железы; 3 — проток железы
У личинок клопа-солдатика (Pyrrhocoris apterus L.), например, каждая из трёх многоклеточных желез, открывающихся на границах четвёртого-шестого тергитов брюшка, имеет все отмеченные структуры, но вместе с тем и каждая входящая в их состав секреторная клетка снабжена собственным внутриклеточным протоком и резервуаром (рис. 63). Пахучие железы взрослых особей этого вида, расположенные на груди, отличаются ещё более сложным строением (рис. 64). Выделяемый ими секрет, как и секрет одноклеточных желез златоглазок, имеет неприятный запах. Выпячивающиеся из сочленовной мембраны между шестым и седьмым тергитом брюшка ароматические железы пчёл издают слабый фруктовый запах, а некоторые клопы пахнут ванилью или земляникой.

Рисунок 64. Пахучая железа имаго клопа-соддатика (по Шванвичу, 1949):
1 — железа; 2 — резервуар; 3 — запирательный клапан; 4 — выводная камера
Весьма своеобразны железы, расположенные на конце брюшка жука-бомбардира Brachinus sp. (рис. 65). В их камерах содержатся фенолы, пероксид водорода и каталаза. При взаимодействии двух последних соединений выделяется кислород, окисляющий фенолы до хинонов. Внешнее проявление этих реакций выражается в том, что сильная струя разогретых хинонов и воды направляется жуком на преследователя. Столь же направлен полет капель жидкого секрета или тончайших, застывающих на воздухе нитей из головного отростка носатых солдат у термитов или муравьиной кислоты, выбрызгиваемой гусеницами гарпии (Cerura vinula L.) из объёмистых желёз у основания передних ног.

Рисунок 65. Отпугивающая железа жука-бомбардира (по Romoser, 1981):
1 — тело железы; 2 — резервуар; 3 — выводная камера
Восковые и лаковые железы червецов и щитовок, белокрылок, тлей, гусениц некоторых бабочек и личинок перепончатокрылых образованы либо отдельными клетками гиподермы, либо небольшими скоплениями клеток (рис. 66).
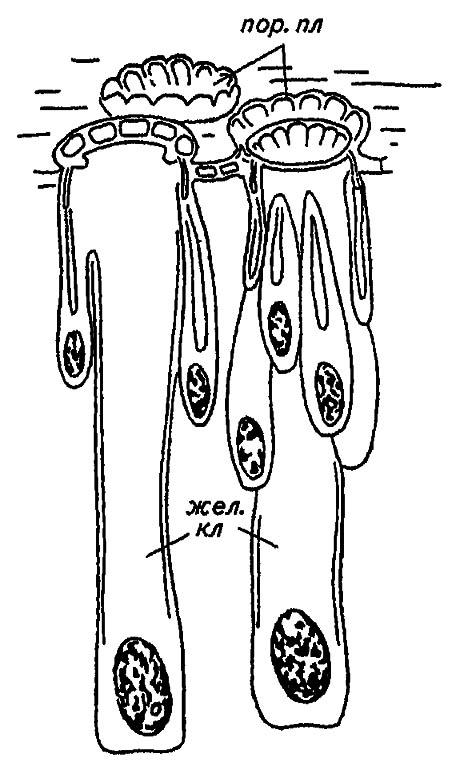
Рисунок 66. Восковые железы червецов (по Шванвичу, 1949, с изменениями):
пор. пл — поровая пластинка; жел. кл — железистые клетки
Выделяя воск сквозь тончайшие поры кутикулы, они придают ему вид ажурных пластинок и нитей, похожих на скульптуру покровов или кристаллы. Эта выделения настолько характерны и специфичны, что позволяют с точностью до вида определить секретирующих их насекомых (рис. 67). У шмелей, пчёл и ос, использующих воск для постройки сотов, соответствующие железы помещаются в сочленениях стернитов брюшного отдела. Формирующиеся в них восковые «таблетки» переносятся к жвалам и затем используются для лепки.
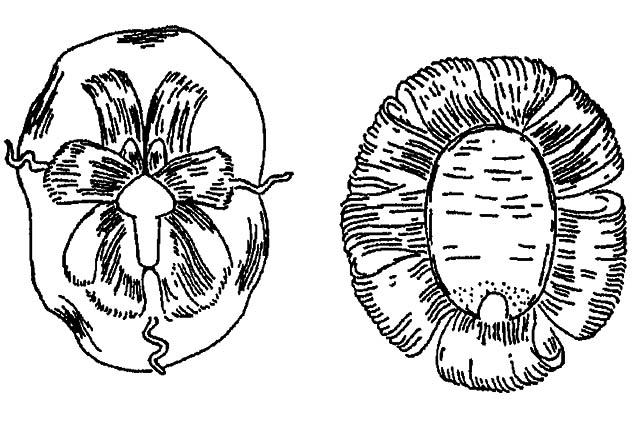
Рисунок 67. Скульптура восковых выделений пупария у белокрылок (по Шванвичу, 1949)
Многочисленные прядильные железы на лапках эмбий образованы секретирующим синцитием, центральным резервуаром и выводным протоком (рис. 68).

Рисунок 68. Прядильные железы эмбий (по Шванвичу, 1949):
А — передняя нога с железами на первом членике лапки; Б — разрез через отдельную железу
Продуцируемый ими шёлк используется для постройки коридоров и галерей в сыпучем грунте, песке или иных субстратах, постоянно загрязняющих их постройки. Особенно оригинально используют шёлкопрядение тропические муравьи Oecophylla. Располагаясь в ряд на краю листа, взрослые особи держат в своих мандибулах выделяющих шёлковые нити личинок. Прочными нитями шёлка они «сшивают» соседние листья по краям в компактные гнезда (рис. 69).

Рисунок 69. Муравьи Oecophylla, строящие гнездо из листьев с использованием своих личинок (по Шванвичу, 1949)
Аллотрофические железы известны у медоносных пчёл, рабочие особи которых выкармливают маточным молочком развивающихся личинок и будущую «царицу» — матку. От режима кормления зависит дальнейшая судьба личинок. Вскармливание в течение двух первых дней гарантирует нормальное развитие рабочих особей. Если личинок кормят до начала метаморфоза, то из образовавшейся крупной куколки формируется матка.
У мирмекофильных, то есть обитающих в муравейниках, жуков семейства Paussidae одноклеточные аллотрофические железы присутствуют на надкрыльях, голове и на сложно устроенных антеннах (рис.70). Выделяя густой секрет, привлекающий муравьёв, жуки избегают участи случайных посетителей муравейника, сразу уничтожаемых. Сходные железы имеются у термитофильных (обитающих в термитниках) жуков-стафилинов, чешуйниц и некоторых других насекомых.

Рисунок 70. Аллотрофические железы на антеннах мирмекофильных жуков Paussidae (по Шванвичу, 1949):
А — левая антенна; Б — антенна, частично вскрытая; жел. кл — железистые клетки; пр — протоки
Гиподермальные железы выделяют феромоны — вещества, привлекающие полового партнёра, других особей своего вида или отмечающие путь, по которому движутся фуражирующие термиты и муравьи. Нередко эти железы весьма сложно устроены; здесь отметим лишь андроконии — волоски и чешуйки на крыльях бабочек, издающие различаемые человеком слабые запахи (рис. 71).

Рисунок 71. Андроконии самца бабочки-голубянки (по Шванвичу, 1949):
жел. кл — железистые клетки; крл. ч — крыловая чешуйка

