Окраска тела
Многие насекомые окрашены скромно и поэтому малозаметны. Другие демонстрируют поразительные интенсивность и разнообразие цветов, которым трудно найти аналогии в живой природе. Например, крылья крупных тропических бабочек рода Morpho сверкают, как полированный голубой металл. Ярко вспыхивая на солнце, они издали могут быть приняты за световые сигналы самолёта, а в сумерках — за мерцание звёзд.
Назначение окраски многообразно. Различают яркую, бросающуюся в глаза апосематическую окраску, предостерегающую потенциальных врагов о возможной ядовитости жертвы. Напротив, известны примеры криптической, гармонирующей с фоном и маскирующей окраски. Характерные цвета часто позволяют опознавать особей своего вида и полового партнёра. Не менее существенно значение окраски для терморегуляции.
При сочетании контрастирующих цветов формируется рисунок тела, состоящий, как правило, из отдельных пятен и полос. Понятно, что окраска тела и его рисунок определяются обычными оптическими эффектами. При пигментной окраске те или иные красители поглощают или отражают световые волны определённого диапазона. Структурная окраска вообще не требует пигментов. Она возникает на основе рассеивания световых лучей, их интерференции или в более редких случаях дифракции тончайшими структурами кутикулы. Однако в сочетании с пигментной окраской она может резко усиливаться и отличаться совершенно неожиданными проявлениями.
В зависимости от локализации пигментов различают весьма стойкую кутикулярную окраску, сохраняющуюся даже в отпечатках ископаемых насекомых, менее стойкую гиподермальную, красящие агенты которой сосредоточиваются в клетках гиподермы, и наконец субгиподермальную, то есть просвечивающую сквозь прозрачные покровы окраску жирового тела, гемолимфы или содержимого кишечника. По формальному критерию все пигменты тела насекомых делят на безазотистые, обычно заимствуемые фитофагами из растений, и азотсодержащие, синтезируемые самими насекомыми. Исключения составляют некоторые безазотистые субгиподермальные афины тлей, неизвестные в растениях, и азотсодержащие тетрапиррольные пигменты, синтез которых тесно связан с метаболизмом поглощённого хлорофилла.
Самые обычные и широко распространённые пигменты насекомых и животных других групп — меланины. Образованные полимеризованными индольными или хиноновыми кольцами, они обычно отлагаются в экзокутикуле, придавая ей тёмную окраску, особенно интенсивную в местах повышенной склеротизации. Однако у палочника Carausius morosus Вr. гранулы этого пигмента заполняют клетки гиподермы и, перемещаясь внутри них, содействуют изменению цвета тела и терморегуляции. Полагают, что образование и последующее отложение меланинов в периодически сбрасываемой кутикуле избавляют организм от токсичных фенольных соединений, вырабатывающихся при склеротизации покровов.
В отличие от меланинов каротиноиды относятся к обычным пигментам растений и воспринимаются питающимися на них насекомыми. Представляя собой непредельные высокомолекулярные углеводороды, каротиноиды обычно связываются с белками и обеспечивают разнообразие окрасок от желтоватых покровов у зрелых особей пустынной саранчи (Schistocerca gregaria Forsk.) до ярко-красных у божьих коровок (Coccinellidae) и красноклопов (Pyrrhocoridae). В сочетании с голубым мезобиливердином из группы тетрапиррольных пигментов они придают зелёную окраску покровам многих прямокрылых.
При питании колорадского жука (Leptinotarsa decemlineata Say.) на картофеле каротиноиды переходят в его гемолимфу и отлагаются в надкрыльях. Нападающий на него хищный клоп Perilius bioculatus Sauss, высасывая окрашенную гемолимфу жертвы, приобретает жёлтую и красную окраску своих собственных покровов. Сходные цвета некоторых полужесткокрылых и бабочек-бархатниц (Satyriadae) обусловливаются флавоноидами, также заимствуемыми ими из растений. У равнокрылых отмечены антракоиноны и близкие к ним афины, которые, по-видимому, не встречаются ни у кого более. Эти вещества широко распространены в мире растений, но насекомые синтезируют их самостоятельно, правда, из тех же исходных продуктов. Некоторые пигменты такого рода флуоресцируют в ультрафиолетовом свете и используются человеком с давних времён. В частности, вырабатываемая кошенилью карминовая кислота не выцветает столетиями, и окрашенные ею средневековые миниатюры до сих пор сохраняют исходную яркость и сочность цвета.
Обычные у чешуекрылых и перепончатокрылых птерины — обогащённые азотом гетероциклические соединения птеридина — определяют жёлтую, белую или красную окраску этих насекомых. Являясь конечными продуктами метаболизма, они, однако, не родственны мочевой кислоте, как полагали ранее. Нередко они сосредоточиваются в глазах насекомого и флуоресцируют, но выделенные из тела дрозофилы пигменты имеют необычный для соединений этой группы голубой цвет.
Из аминокислоты триптофана при участии ДОФА синтезируются оммохромы. Концентрируясь в покровах и фасеточных глазах многих насекомых, они придают глазам коричневый, жёлтый или красный цвет. Яркие крылья бабочек-нимфалид и розовая окраска тела преимагинальных стадий развития пустынной саранчи связывают именно с этими пигментами.
Тетрапиррольные соединения, относящиеся к пигментам, входят также в состав хлорофилла, гемоглобина, цитохромов. Они, видимо, синтезируются из глицина. Один из пигментов этой группы — биливердин определяет зелёный цвет покровов богомолов, некоторых саранчовых и палочников. Способный с лёгкостью окисляться и восстанавливаться, он обеспечивает приспособительное изменение окраски их тела.
С оптическими эффектами рассеивания, интерференции и дифракции световых волн на тонких структурах поверхности тела насекомых связаны интенсивность, разнообразие, изменчивость и радужность их окраски. При этом, если оптически активные структуры кутикулы относительно велики по сравнению с длиной волн света и распределены нерегулярно, возникают снежно-белые цвета. Если расстояния между регулярно расположенными структурами сравнимы с длиной световых волн, то возникает интерференция, приводящая к усилению определённых цветов, меняющихся при изменении угла зрения.
Тончайшие и прозрачные крылья златоглазок и мелких подёнок переливаются всеми цветами радуги, усиливающимися при рассматривании на чёрном фоне. Особенно сильное впечатление производит интерференционная окраска оптических чешуек крыльев бабочек тропического рода Urania. Их чёрный основной фон сияет ярко-зелёным светом, к которому добавляются золотисто-жёлтые, красные, синие, фиолетовые и пурпурные отблески. Рассредоточенные среди оптически нейтральных чешуек, они придают крыльям в целом характерный шагреневый перелив.
Сходные оптические эффекты возникают на крыльях многих парусников и голубянок. У южноамериканских бабочек рода Morpho и многих тропических нимфалид в окраске крыльев преобладают интенсивные синие и голубые тона, по которым переливаются широкие ярко-зелёные и голубые волны. Иную природу имеют голубые пятна на крыльях некоторых бабочек и радужная окраска покровов ряда жуков. Такая окраска образуется на тончайших дифракционных решётках кутикулярных структур и сохраняется даже на искусственных слепках с поверхности, сделанных из коллодия. Серебристый блеск крыльев широко распространённых бабочек-перламутровок определяется сложными сочетаниями пигментной окраски с результатами интерференционных и дифракционных явлений. В целом дифракционная окраска, по-видимому, редка в мире насекомых, и почти все металлически-блестящие цвета тела жуков-златок и листоедов обусловлены интерференцией, усиленной тёмной пигментной основой.
Нередко контрастирующие цвета окраски образуют рисунок. На крыльях бабочки-перламутровки элементы рисунка — тёмные меланиновые пятна на жёлтом фоне — при нарушении синтеза меланина остаются белыми. Следовательно, при одном и том же рисунке окраска может быть различной.
Особенно сложны и прихотливы рисунки на крыльях бабочек; значительно менее развит рисунок покровов жуков, что не связано с окраской (ведь многие ярко окрашенные листоеды и златки вообще не имеют рисунка). Среди остальных насекомых рисунок имеется у прямокрылых, клопов, ручейников, скорпионниц и цикад.
Несмотря на разнообразие и изменчивость отдельных компонентов рисунка крыла, например у бабочек-нимфалид, можно выделить его общую схему и прототип. По мнению Б. Н. Шванвича, прототип рисунка слагается из пяти систем поперечных полос (рис. 72): первой — трёх узких экстерн вдоль края крыла; второй, следующей за ними, — глазчатых пятен со зрачками; третьей — двух дискальных полос близ дискальной ячейки крыла и базальной полосы у его основания; четвёртой — двух медиальных, или срединных, полос; пятой системы — умбр (теней) с расплывчатыми контурами, которые заполняют пространства между поперечными полосами.

Рисунок 72. Прототип рисунка крыльев бабочек семейства нимфалид (А) и парусников (Б) (по Шванвичу, 1949):
экс — экстерны; глч. п — глазчатые пятна; мед, баз, дис — соответственно медиальные, базальные, дискальные полосы; ум — умбры
Преобразования этого общего прототипа сводятся к выпадению или усилению отдельных частей рисунка, к изгибаниям и перемещениям полос, образованию комплексов, обмену местами и наложению некоторых полос друг на друга. При этом оказывается, что наименее постоянна вторая экстерна, а наиболее — третья; из системы глазчатых пятен особенно прочны второе пятно заднего крыла и пятое пятно переднего, причём глазчатые пятна отличаются особой «активностью». Сближаясь, но не соприкасаясь с другими элементами рисунка, они вызывают их смещения и изгибания. В системе дискальных полос первая резко ограничена снаружи и расплывчата у внутреннего края, тогда как вторая, наоборот, размыта с внешней стороны и имеет чёткий контур с внутренней.
Существенно, что выявленные преобразования рисунка регистрируются в разных группах чешуекрылых, не связанных друг с другом непосредственным родством. В этих параллельно осуществляющихся тенденциях очевидно проявление некоторых общих закономерностей изменения рисунка, известных нам лишь частично.
Не всегда понятны функциональный и приспособительный смысл тех или иных окрасок и рисунков, а также биологическая целесообразность их преобразований. С несколько большей определённостью можно судить о значении сезонных и возрастных изменений: необратимых, связанных с синтезом, накоплением и распадом пигментов, и обратимых, вызванных смещением пигментных гранул или специальных пигментных клеток — меланофоров.
Особенно наглядна хроматическая адаптация прямокрылых, обитающих в африканской саванне. После нередких здесь пожаров покровы насекомых в считанные дни темнеют, сближаясь по цвету с преобладающим фоном выжженной травы и кустарников. Приспособительное значение этих изменений окраски очевидно.
Менее понятно регулярное изменение окраски широко распространённых бабочек-голубянок или огородных клопов. В первом случае весенние и осенние формы, например Lycoenaphylaens daimio Hb., отличаются ярко-оранжевым фоном крыльев, а летние формы сравнительно темные.
Зимующие клопы Eurydema ornatum L. имеют на теле тёмные пятна на более светлом фоне, который меняет цвет у летних и осенних форм (рис. 73). Примечательно, что обширные тёмные пятна на брюшке клопа Oncopeltus fasciatus L. формируются лишь при комнатной температуре и исчезают при повышении её до 25–27 °С. Закрашивание глаз кобылки Oedipoda coerulescens L. непрозрачным лаком лишает этих насекомых способности приспосабливаться к цвету почвы. Очевидно, что внешние стимулы, воздействуя на рецепторы и нервные центры, активизируют эндокринные железы, определяющие характер и ход пигментации посредством выделяемых ими гормонов. Изменение окраски возможно и без участия внешних стимулов, как, например, у гусениц бабочки гарпии Cerura vinula L., краснеющих перед окукливанием. Наблюдаемое здесь отложение оммохромов — лишь побочный результат действия линочного гормона.

Рисунок 73. Сезонные изменения окраски клопа Eurydema ornatum L. (по Шванвичу, 1949):
А, Б, В, Г — соответственно осенняя, зимняя, весенняя и летняя окраски
Приведённые примеры демонстрируют индивидуальные приспособления, возможности отдельных особей к изменению цвета тела в течение жизни. Эти изменения непрямое следствие отбора, как при индустриальном меланизме берёзовых пядениц (Biston betularia L.), среди которых выживаемость более тёмных форм существенно выше (на 30 %), чем светлоокрашенных. Последние гораздо заметнее для хищников на загрязнённом дымом и копотью фоне (рис. 74).

Рисунок 74. Индустриальный меланизм берёзовой пяденицы Biston betularia L. (по Фарбу, 1976)
Более очевиден адаптивный характер обратимых и относительно быстрых изменений окраски тела насекомых. У палочника Carausius morosus Вr. клетки гиподермы включают гранулы четырёх пигментов: зелёные и жёлтые распределены равномерно по цитоплазме, а коричневые и оранжевые способны перемещаться либо к основанию клеток, и тогда тело становится более светлым, либо к их вершинам — и тело темнеет. Движения гранул и изменения окраски тела определяются множеством стимулов: изменением цвета фона, температуры, осмотического давления гемолимфы, влажности, механическими воздействиями и зрительными восприятиями — и вместе с тем подвержены суточному ритму: днем палочник окрашен светлее, чем ночью. Сходные изменения характерны для некоторых богомолов и, по-видимому, гусениц бабочек. Они содействуют маскировке насекомых, которые не способны к активной самозащите.
Разительно сходство гусениц бабочек-пядениц и палочников с высохшими сучьями или палочника странствующего листа Phyllium и бабочки Callima с листьями растений (рис. 75). Это сходство может ввести в заблуждение зоркого хищника.
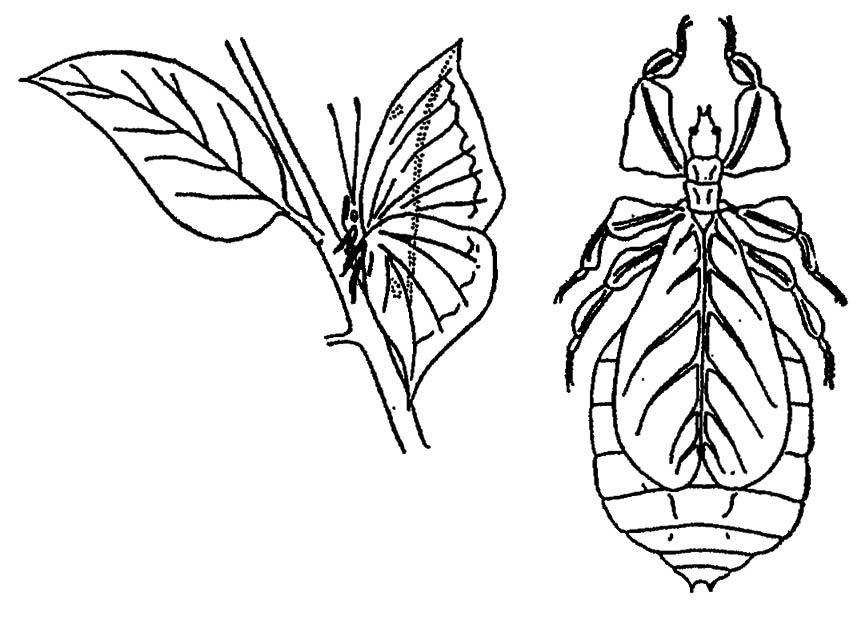
Рисунок 75. Листовидная форма тела бабочки Callima и палочника Phyllium (по Шванвичу, 1949)
Многие ядовитые и агрессивные насекомые, которые могут стать случайными жертвами неопытных хищников, вырабатывают общие признаки, символизирующие опасность. Таковы ярко-окрашенные жуки-нарывники, бабочки с ядовитой гемолимфой, осы и пчелы, демонстрирующие апосематическую (угрожающую) окраску. Вместе с тем эти хорошо защищённые насекомые нередко становятся образцами для подражания, своеобразными моделями для других, вполне съедобных и беззащитных форм. Обладая угрожающей внешностью, они, естественно, повышают свои шансы на выживание, однако данный тип мимикрии, открытый Бейтсом, имеет одно существенное ограничение. Дело в том, что если подражатели становятся многочисленнее своих моделей, то последние станут чаще подвергаться нападению хищников, уже приобретших опыт общения с пугающими, но незащищёнными формами.
Особенно интересный пример бейтсовской мимикрии отмечен у южноафриканского парусника (Papilio dardanus L.), желто-окрашенные самцы и самки которого обычно обитают на Коморских островах, Мадагаскаре и в Сомали. Однако в Эфиопии и других регионах самки парусника становятся похожими на различных несъедобных бабочек семейства Danaidae, а самцы остаются неизменными (рис. 76). Таким образом, в пределах ареала однообразным самцам противопоставляется множество разнообразных самок, подражающих разным моделям и отличающихся от них лишь малозаметными, но характерными для парусников отростками задних крыльев. При скрещивании всех этих форм наблюдается обычное моногибридное расщепление, жёстко сцепленное с полом. Следовательно, каждая миметическая форма контролируется, по существу, одним геном (супергеном), локализованным на половой хромосоме самок. Учитывая, что исходные, немиметирующие самки обитают на границе ареала, можно объяснить их многообразие сочетанием дизруптивного и движущего, но постоянно меняющего своё направление отбора.

Рисунок 76. Мимикрия Papilio dardanus (по Эрлиху, Холму, 1966):
А — самец; Б — самки разных миметических форм; В — представители семейства, являющиеся моделями
В заключение отметим, что среди разнообразия форм насекомых выделяются некоторое постоянство и единство, связанные с общностью их происхождения, образа жизни и условий существования, а также с общностью системных объектов.
Каждый объект как система, образованная некоторым числом взаимодействующих элементов, в соответствии с общей теорией систем формируется за счёт либо изменения самих элементов, либо их числа и существующих между ними взаимоотношений и связей, либо всех указанных изменений. Нетрудно убедиться, что живые объекты создаются в природе ограниченным набором способов, отвечающим числу неповторяемых сочетаний элементов, отношений и числа элементов.
Системная общность рассматриваемых нами объектов выражается, например, в разительном сходстве жабр у личинок подёнок и стрекоз с зачатками крыльев, членистых антенн с церками, мощных форцепсов с челюстями. Сходство насекомых или отдельных частей их тела может быть генетическим, конвергентным и системным.

