Разнообразие ротовых аппаратов
Своеобразие грызуще-лижущего ротового аппарата перепончатокрылых (рис. 14) определилось в эволюционных взаимодействиях с опыляемыми ими растениями. Сохранив все основные структуры прототипа (грызущего ротового аппарата), он сформировался за счёт редукции одних компонентов или усиленного развития других. При этом в наименьшей степени изменились верхняя губа и мандибулы. С помощью мандибул, принимающих вид узких лопаточек, насекомые строят гнезда, а в покое мандибулы плотно прижимают сложенные под головой максиллы и нижнюю губу. Последние преобразуются в узкие лопасти, слагающиеся в удлинённый хоботок для всасывания нектара.
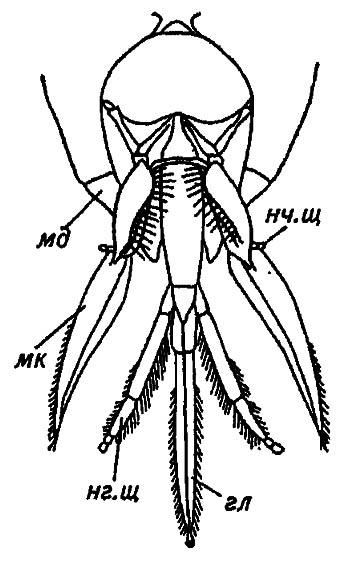
Рисунок 14. Грызуще-лижущий ротовой аппарат перепончатокрылых (по Gillot, 1980)
А — верхние челюсти (мандибулы): мд. м — мандибулярные мыщелки; м. к — молярный (коренной) край; р. к — режущий край мандибулы; Б — нижние челюсти (максиллы): кар — кардо (основной членик); ст — стипес (стволик); г — галеа; лац — лациния; нч. щ — нижнечелюстной щупик; В — нижняя губа: пб — подбородок (ментум); пм — постментум; прм — прементум; гл — глоссы; пгл — параглоссы; нг. щ — нижнегубной щупик
Кардо — основной членик максилл, представляет собой стержень, сочленённый с краем головной капсулы, с сильно изменённым стволиком, несущим рудимент нижнечелюстного щупика. Особенно сильно развиты галеа, тогда как лацинии образуют небольшие эластичные створки у основания хоботка. Среди компонентов нижней губы можно выделить субментум, образующий вилочку (lorum), между ветвями которой помещён треугольный подбородок, и прементум — вытянутую пластинку с причленёнными у её вершины длинными нижнегубными щупиками, короткими параглоссами и тонкими, сросшимися друг с другом глоссами.
Таким образом, нижние челюсти и нижняя губа, сохранив все основные структуры грызущего ротового аппарата, сформировали хоботок, причём его дном стал прементум со своими придатками, а боковыми стенками — стволики и галеа максилл. Сгибаясь по продольному шву и налегая друг на друга краями, они замыкаются в узкую трубку, открытую с обоих концов у вершины и основания. Однако отверстие у основания, служащее для кормления личинок маточным молочком, способно плотно замыкаться лациниями в момент всасывания нектара. Вершинное отверстие хоботка остаётся незамкнутым, и из него выдаются сросшиеся глоссы. Густо покрытые щетинками, они заканчиваются своеобразной маленькой ложечкой и, вибрируя при погружении в нектарник, содействуют продвижению нектара вверх, к ротовому отверстию. Наряду с этим продвижение жидкой пищи по хоботку обеспечивается действием глоточного насоса.
Расположенные на одном уровне сочленения нижних челюстей и нижней губы (лабиомаксиллярные сочленения) позволяют хоботку складываться в два колена и прижиматься мандибулами к голове. Вместе с тем, приведённый в рабочее состояние, развёрнутый хоботок благодаря гибкости околоротовой мембраны и вышеназванным сочленениям может сильно выдвигаться вперёд и проникать в узкие глубокие венчики цветков. Среди обычных для нашей фауны перепончатокрылых-опылителей особенно длинным хоботком обладают шмели, и это позволяет им кормиться даже на культурных сортах клевера ползучего (красного), глубокие нектарники которого недоступны для пчёл.
Особенно радикальные преобразования связаны с оформлением ротового аппарата чешуекрылых (рис. 15). Его основу составляют сильно вытянутые галеа. Свёрнутые в покое под головой плотной спиралью, они способны стремительно развернуться в длинный и тонкий хоботок, проникающий в глубокие венчики цветков. У некоторых бражников галеа превосходят длину тела.

Рисунок 15. Сосущий ротовой аппарат чешуекрылых (по Шванвичу, 1949, с изменениями)
А — верхние челюсти (мандибулы): мд. м — мандибулярные мыщелки; м. к — молярный (коренной) край; р. к — режущий край мандибулы; Б — нижние челюсти (максиллы): кар — кардо (основной членик); ст — стипес (стволик); г — галеа; лац — лациния; нч. щ — нижнечелюстной щупик; В — нижняя губа: пб — подбородок (ментум); пм — постментум; прм — прементум; гл — глоссы; пгл — параглоссы; нг. щ — нижнегубной щупик
Наряду с ними у многих бабочек сохраняются рудиментарные нижнечелюстные щупики, тогда как щупики нижней губы обычно хорошо развиты, несмотря на значительное сокращение других её структур. Мандибулы незаметны, а редуцированная верхняя губа лишь прикрывает сверху основание хоботка. Отметим, что у примитивных форм, например у зубатых молей, мандибулы вполне развиты, как и другие компоненты грызущего ротового аппарата, но галеа сравнительно невелики. Существенно, что это примитивное состояние, близкое к описанной ранее схеме, связано рядом переходов с состоянием высокой специализации, весьма характерным для чешуекрылых; при этом последовательному удлинению галеа сопутствует последовательная редукция других компонентов грызущего ротового аппарата (рис. 16).

Рисунок 16. Последовательные этапы формирования хоботка у чешуекрылых (по Шванвичу, 1949):
А — исходное состояние у Micropterygidae; Б — удлинение галеа у Mnemonia auricana; В — хоботок Pronuba yuccasella; Г — хоботок Synanthedon exitiosa.
Несмотря на внешнюю простоту строения, сосущий хоботок бабочек весьма совершенен. Обе длинные и гибкие галеа соединены друг с другом по всей длине скользящим сочленением и плотно прилегают друг к другу свободными краями. Формирующаяся при этом трубка служит для проведения нектара, засасываемого мощным насосом, расположенным внутри головы и представленным видоизменёнными стволиками максилл. Вместе с тем галеа имеют собственные полости, сообщающиеся с полостью тела. С силой нагнетая в них полостную жидкость, бабочка быстро разворачивает тугую спираль хоботка, приводя его в деятельное состояние. Закончив питание, она ослабляет давление, и хоботок снова скручивается спиралью, как часовая пружина.
Итак, мы остановились на ротовых аппаратах двух типов: грызуще-лижущем перепончатокрылых и сосущем — чешуекрылых. Различаясь по набору структур и тенденциям эволюции, они тем не менее выполняют сходные функции, сводящиеся к всасыванию жидкой пищи. Рассматривая колюще-сосущие ротовые аппараты комаров и клопов, мы увидим иные преобразования исходного грызущего ротового аппарата, которые наряду со всасыванием жидкой пищи обеспечивают прокалывание ограничивающих её оболочек.

Рисунок 17. Колюще-сосущий ротовой аппарат полужесткокрылых (по Шванвичу, 1949)
гл — глаза; ант — антенны; глз — глазки; в. г — верхняя губа; мд — мандибулы; мк — максиллы; н. г — нижняя губа; т. ш — теменной шов; л. ш — лобные швы; л — лоб; нал — наличник; щ — щёки; пщ — подщёки; зщ — защёки; зт — затылок; ззт — заднезатыпок; з. о — затылочное отверстие; р. от — ротовое отверстие; т. я — тенториальные ямки; гпф — гипофаринкс; з. ш — затылочный шов; ззт. ш — заднезатылочный шов; ш.скл — шейные склериты; сл. п — отверстие слюнного протока
Конструктивной основой колюще-сосущего ротового аппарата клопов и близких к ним равнокрылых служат четыре тонких стилета (рис. 17, 18). Соответствующие мандибулам и максиллам, они заключены в членистую нижнюю губу, выполняющую роль ножен, и в момент прокалывания покровов способны выдвигаться из неё (рис. 19).

Рисунок 18. Поперечный срез через стилеты ротового аппарата клопа (по Шванвичу, 1949)
гл — глаза; ант — антенны; глз — глазки; в. г — верхняя губа; мд — мандибулы; мк — максиллы; н. г — нижняя губа; т. ш — теменной шов; л. ш — лобные швы; л — лоб; нал — наличник; щ — щёки; пщ — подщёки; зщ — защёки; зт — затылок; ззт — заднезатыпок; з. о — затылочное отверстие; р. от — ротовое отверстие; т. я — тенториальные ямки; гпф — гипофаринкс; з. ш — затылочный шов; ззт. ш — заднезатылочный шов; ш.скл — шейные склериты; сл. п — отверстие слюнного протока

Рисунок 19. Последовательные акты внедрения стилетов ротового аппарата клопов в ткани питающего организма (по Weber, 1966)
гл — глаза; ант — антенны; глз — глазки; в. г — верхняя губа; мд — мандибулы; мк — максиллы; н. г — нижняя губа; т. ш — теменной шов; л. ш — лобные швы; л — лоб; нал — наличник; щ — щёки; пщ — подщёки; зщ — защёки; зт — затылок; ззт — заднезатыпок; з. о — затылочное отверстие; р. от — ротовое отверстие; т. я — тенториальные ямки; гпф — гипофаринкс; з. ш — затылочный шов; ззт. ш — заднезатылочный шов; ш.скл — шейные склериты; сл. п — отверстие слюнного протока
При этом сначала продвигается вперёд одна из мандибул, затем — вторая, и лишь после того, как они плотно укрепятся в тканях зазубренными краями, одним быстрым движением выдвигаются обе максиллы. Далее все операции повторяются заново, и так шаг за шагом клоп вводит стилеты в ткани питающего организма.
У некоторых форм стилеты намного длиннее тела и в покое свёрнуты широкой петлёй или спиралью внутри головы. Для того чтобы ввести их целиком, насекомое после каждого внедрения стилетов плотно сжимает их нижней губой и, отводя голову назад, постепенно разворачивает спирали и петли. При этом стилеты не выдёргиваются из ранки и при следующем внедрении проникают на ещё большую глубину. Одновременно через канал, образованный противолежащими желобками на внутренней поверхности максилл, в ранку вводится слюна, обладающая протеолитическими, а у паразитов и анастезирующими свойствами и способностью разжижать кровь животных и соки растений. Именно поэтому прокол может не вызвать болезненных ощущений, которые появляются лишь впоследствии. Наряду со слюнным каналом аналогичные желобки на максиллах образует канал для всасывания жидкой пищи (см. рис. 17).
Понятно, что прокалывание жёстких покровов требует значительного усилия, которое трудно развить мелким, легковесным цикадкам и тлям. Однако опистогнатическая постановка головы позволяет смещать стилеты к центру тяжести тела, что облегчает исполнение этой задачи. Добавим, что максиллярные стилеты, по-видимому, соответствуют лациниям; другие компоненты ротового аппарата, а именно верхняя губа и рудиментарный гипофаринкс, не участвуют в проколе.
У низших двукрылых, например комаров, в отличие от полужесткокрылых и равнокрылых колюще-сосущий аппарат образован шестью стилетами (рис. 20).

Рисунок 20. Колюще-сосущий ротовой аппарат комаров (по Шванвичу, 1949)
гл — глаза; ант — антенны; глз — глазки; в. г — верхняя губа; мд — мандибулы; мк — максиллы; н. г — нижняя губа; т. ш — теменной шов; л. ш — лобные швы; л — лоб; нал — наличник; щ — щёки; пщ — подщёки; зщ — защёки; зт — затылок; ззт — заднезатыпок; з. о — затылочное отверстие; р. от — ротовое отверстие; т. я — тенториальные ямки; гпф — гипофаринкс; з. ш — затылочный шов; ззт. ш — заднезатылочный шов; ш.скл — шейные склериты; сл. п — отверстие слюнного протока
Кроме тонких мандибул и максилл в прокалывании участвуют удлинённые верхняя губа и гипофаринкс. Первая напоминает косо срезанную иглу шприца и особенно прочна. Являясь опорой для других стилетов, она вместе с тем предназначена для проведения пищи, тогда как более тонкий гипофаринкс — для проведения слюны. Осуществляя прокол, комар прочно цепляется за покровы коготками лапок и, надавливая на хоботок, пропиливает кожу зазубренными максиллами. Последние, видимо, гомологичны галеа и вместе с другими стилетами окружены, как футляром, нечленистой нижней губой (рис. 21).

Рисунок 21. Поперечный срез через хоботок комара (по Weber, 1966)
гл — глаза; ант — антенны; глз — глазки; в. г — верхняя губа; мд — мандибулы; мк — максиллы; н. г — нижняя губа; т. ш — теменной шов; л. ш — лобные швы; л — лоб; нал — наличник; щ — щёки; пщ — подщёки; зщ — защёки; зт — затылок; ззт — заднезатыпок; з. о — затылочное отверстие; р. от — ротовое отверстие; т. я — тенториальные ямки; гпф — гипофаринкс; з. ш — затылочный шов; ззт. ш — заднезатылочный шов; ш.скл — шейные склериты; сл. п — отверстие слюнного протока
Перейдём к рассмотрению наиболее своеобразного и универсального ротового аппарата насекомых — мускоидного, лижущего, или фильтрующего, хоботка высших мух (рис. 22).

Рисунок 22. Мускоидный (лижущий) ротовой аппарат высших двукрылых (по Шванвичу 1949):
в. г — верхняя губа; мк. щ — максиллярный щупик; рст — рострум; гст — гаустеллум; лаб — лабеллумы
Представленный гипертрофированной нижней губой, он сохранил видоизменённые максиллы, гипофаринкс и верхнюю губу, но полностью утратил мандибулы. Основание хоботка, или его рострум, образовано разросшимися краями головной капсулы, наличником, нижнечелюстным щупиком и максиллярным склеритом, служащим для складывания хоботка. Верхняя губа и гипофаринкс включены в гаустеллум. По-видимому, он соответствует прементуму, имеющему видоизменённые нижнегубные щупики — лабеллумы. И рострум, и гаустеллум, и лабеллумы подвижно соединены друг с другом. Складываясь двумя коленами под головой, в недеятельном состоянии они почти незаметны.
При разворачивании хоботка особой подвижностью отличаются лабеллумы, имеющие вид двух полукруглых присосок с расположенным между ними по центру отверстием для приёма пищи (рис. 23).

Рисунок 23. Разворачивающиеся лабеллумы ротового аппарата высших двукрылых в положениях покоя (А), фильтрации жидкой пищи (Б), обскрёбывания субстратов (В):
з — зубы; пс — псевдотрахеи; м — гибкая мембрана; мц — мышцы; скл — склериты
К этому отверстию подходят погруженные под поверхность лабеллумов псевдотрахеи — тонкие трубочки с мелкими порами, укреплённые полукружными склеритами. Через поры отфильтровывается полужидкая пища. Проникая в псевдотрахеи, она проходит через центральное отверстие в гаустеллум и рострум и в конце концов достигает глотки в голове насекомого. На границе центрального отверстия, между подходящими к нему псевдотрахеями, расположены крепкие зубцы. При выворачивании лабеллумов они обнажаются и служат для дробления твёрдых субстратов, а при складывании погружаются внутрь.
Таким образом, наряду с фильтрацией полужидкой пищи и всасыванием воды мускоидный хоботок допускает питание твёрдой пищей, которая обычно разжижается выделяемой слюной. Благодаря мягкости всех компонентов и эластичности мембраны лабеллумов хоботок высших мух способен охватывать мелкие частички пищи, плотно прижиматься к субстратам и проникать в довольно узкие щели. Расположенные внутри него склериты и мышцы обеспечивают весьма разнообразные движения лабеллумов, складывание и разворачивание хоботка, а также всасывание пищи.
Широкое разнообразие функций мускоидного ротового аппарата обусловливает эволюционный прогресс высших двукрылых. Лишь неспособность к прокалыванию покровов несколько ограничивает набор используемых пищевых субстратов. За счёт незначительных преобразований лабеллумов некоторые представители подотряда становятся кровососами.
Укрупнение и увеличение числа зубцов у пищеприёмного отверстия позволяют жигалкам и мухам цеце просверливать даже толстую кожу млекопитающих: плотно прижимая хоботок и попеременно выворачивая и вворачивая лабеллумы, эти насекомые процарапывают кожу до появления крови (рис. 24).
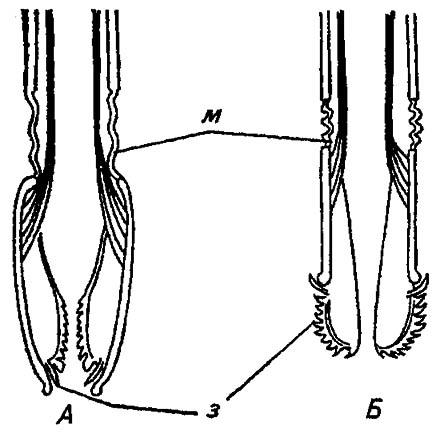
Рисунок 24. Сверлящий ротовой аппарат высших двукрылых (жигалок и мухи цеце, видоизменено по Шванвичу, 1949):
А, Б — соответственно свёрнутый и развёрнутый хоботок. з — зубы; пс — псевдотрахеи; м — гибкая мембрана; мц — мышцы; скл — склериты
Таким образом, в пределах названной группы реализуются почти все возможные для насекомых способы приёма пищи.
Нами рассмотрены лишь некоторые типы ротовых аппаратов, многие их модификации остались за пределами обсуждения. Однако приведённых сведений достаточно для демонстрации эволюционной пластичности отдельных структур и наиболее общих тенденций их преобразования. Соответствующие гомологии отдельных структур ротовых органов насекомых представлены на рисунке 25.

Рисунок 25. Гомологии ротовых органов насекомых:
а — грызуще-лижущие; б — грызущие (ортоптероидные); в — грызущие (ручейников); г — режущие (слепней); д — сосущие (хоботок бабочек); е, ж — колюще-сосущие (клопов и комаров); з — лижущие (мух); 1 — верхняя губа; 2 — мандибулы; 3 — нижняя губа; 4 — максиллы

