Брюшко и его придатки
Брюшко насекомых состоит из обособленных сегментов, которые не имеют развитых ног и локомоторной мускулатуры, сравнительно однообразны и мало видоизменены. Их тергиты и стерниты связаны между собой гибкой плейральной мембраной с немногочисленными склеритами и сегментарно расположенными отверстиями дыхалец — стигм (см. рис. 3).

Рисунок 3. План строения насекомого:
гол — голова; I, II, III — последовательные сегменты грудного отдела; 1-7 — прегенитальные; 8, 9 — генитальные; 10, 11 — постгенитальные сегменты брюшка; крл — крылья; нг — ноги; пол. пр — половые придатки; цер — церки; рот. пр — ротовые придатки; ан — анальное отверстие; ст — стигмы
Слабое развитие плейритов соответствует неразвитости придатков.
У примитивных форм в состав брюшка входят не менее 11 сегментов, но у большинства насекомых некоторые из них, чаще всего самые задние, редуцируются или вворачиваются внутрь тела. Являясь средоточием вегетативных функций и отправлений, брюшко, обычно цилиндрическое или веретеновидное, нередко становится уплощённым или округлым; у некоторых термитов и муравьёв (так называемых медовых бочек), накапливающих в своём теле резервы для вскармливания других членов семьи, брюшко значительно растягивается и по размерам намного превосходит другие отделы тела (рис. 46).

Рисунок 46. Раздутое брюшко муравья Anergates (по Шванвичу, 1949)
У жуков-короедов брюшко сильно укорачивается, приобретая вид усечённого конуса (рис. 47); у певчих цикад и подёнок оно преобразуется соответственно в пустотелый резонатор звуков и своеобразный аэростат. Напомним, что у части перепончатокрылых (у Apocrita) первый сегмент брюшка — проподеум входит в состав груди, а второй (у муравьёв — второй и третий) образует тонкий стебелёк.
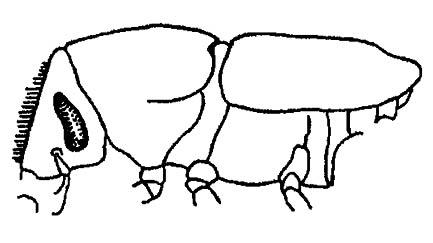
Рисунок 47. Короед-заболонник (по Шванвичу, 1949)
Этот стебелёк обеспечивает подвижность брюшка и возможность нанесения уколов расположенным на его конце жалом или яйцекладом. На восьмом и девятом сегментах сосредоточены половые придатки, поэтому указанные сегменты называются генитальными, а расположенные впереди и позади них — соответственно прегенитальными и постгенитальными (см. рис. 3).
Придатки брюшка у подавляющего большинства насекомых рудиментарны или сильно видоизменены, но у многоножек и близких к ним форм брюшные сегменты имеют развитые конечности, сходные с конечностями грудного отдела.
У представителей отряда бессяжковых (Protura) на трёх первых сегментах брюшка расположены короткие двучлениковые ноги (рис. 48).

Рисунок 48. Рудиментарные брюшные ноги бессяжковых (по Шванвичу, 1949):
тз — тазик; пуз — впячивающийся пузырёк
Их соответствие конечностям грудного отдела и челюстям не вызывает сомнения. У щетинохвосток — истинных первичнобескрылых насекомых — на всех прегенитальных сегментах имеются своеобразные придатки, состоящие из основания, короткого грифелька (stylus) и двух впячивающихся пузырьков (рис. 49), по-видимому, соответствующих двум ветвям конечности ракообразных.

Рисунок 49. Придатки прегенитальных сегментов брюшка щетинохвостки (по Шванвичу, 1949):
стер — стернит; грф — грифелёк; пуз — впячивающиеся пузырьки
У близких к многоножкам ногохвосток за счёт этих образований на первом сегменте брюшка сформировалась брюшная трубка — коллофор (collophore) с раздвоенной подушечкой на конце, а на втором сегменте — своеобразная зацепка — тенакулум (tenaculum). Её зубцы предназначены для фиксации прыгательной вилки (furca), развивающейся на четвёртом сегменте. При сведении зубцов зацепки подогнутая под брюшко вилка освобождается и с силой ударяет по субстрату, подбрасывая тело насекомого высоко вверх. Весьма характерны придатки брюшных сегментов у водных личинок подёнок и вислокрылок; выполняя функции трахейных жабр, они имеют вид перисто-рассечённых лепестков у первых и членистых конечностей у вторых. Возможность гомологизации этих структур с настоящими конечностями не исключена, но требует дополнительных обоснований.
Необходимо упомянуть о брюшных ложноножках гусениц бабочек и ложногусениц примитивных перепончатокрылых. Имея вид коротких мясистых образований, они активно участвуют в движении и обычно представлены крупной субкоксой, коротким тазиком и снабжённой крючьями подошвой (рис. 50).

Рисунок 50. Ложноножка гусеницы (по Шванвичу, 1949):
под — подошва; тз — тазик; субк — субкокса
У гусениц они развиты на третьем—шестом и десятом сегментах, у ложногусениц — на втором—восьмом и десятом. Эти придатки рассматривают либо как редуцированные и видоизменённые конечности брюшных сегментов, либо как новообразования.
Отдельную группу составляют придатки постгенитальных сегментов: парные церки (cercus), непарный хвостовой фш1амент и трахейные жабры личинок равнокрылых стрекоз. Церки развиваются на одиннадцатом сегменте и у многих примитивных насекомых имеют вид длинных членистых нитей, напоминающих щетинковидные антенны. Однако у уховёрток и некоторых вилохвосток они преобразуются в своеобразные клещи — форцепсы. У высших насекомых церки обычно не развиты. Каудальный филамент вместе с церками образует, например, у подёнок три длинные хвостовые нити, но у большинства насекомых он редуцируется. У личинок стрекоз он даёт начало листовидной трахейной жабре и вместе с боковыми жабрами, развившимися на склеритах близ анального отверстия, обеспечивает дыхание этих водных обитателей. Гомологизация данных образований с сегментарными конечностями вряд ли возможна.
Переходя к описанию придатков генитальных сегментов, уместно рассмотреть строение наиболее примитивных насекомых, а именно щетинохвосток, имеющих рудиментарные брюшные ноги. Их основания образованы широкими пластинками — коксоподитами, к заднему краю которых причленены грифельки (см. рис. 49). Однако коксоподиты восьмого и девятого сегментов отличаются тем, что, во-первых, не имеют впячивающихся подушечек; во-вторых, снабжены длинными выростами — гонапофизами, отходящими от передних срединных углов каждой пластинки (рис. 51).

Рисунок 51. Яйцеклад щетинохвосток (по Шванвичу, 1949):
коксп — коксоподиты соответствующих сегментов; грф — грифельки; гон I — первый гонапофиз
Эти парные выросты, близко подходя друг к другу, объединяются в своеобразный яйцеклад, между четырьмя створками которого проскальзывает откладываемое самкой яйцо. Гонапофизы самцов щетинохвосток более короткие (рис. 52), чем у самок; на сочленовной мембране между восьмым и девятым сегментами у самцов формируется непарный копулятивный орган, разделённый на основание — фаллобазу и трубчатый эдеагус. В отличие от гонапофизов он не гомологичен конечностям и представляет собой новообразование.
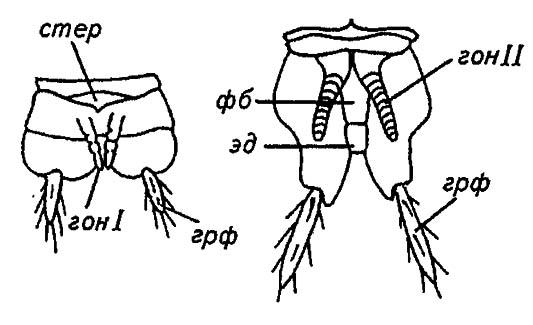
Рисунок 52. Копулятивные придатки самца щетинохвостки (по Шванвичу, 1949):
стер — стернит; гон I и гон II — первые и вторые гонапофизы соответственно; эд — эдеагус; фб — фаллобаза; грф — грифельки
Приведённая схема позволяет понять строение более специализированных половых придатков высших насекомых. В частности, яйцеклад Pterygota включает те же самые структуры, которые, однако, именуются иначе: коксоподиты называются яйцекладными, или основными, пластинками, а гонапофизы — створками яйцеклада. Первая пара створок принадлежит восьмому, а вторая — девятому сегменту. Вместе с тем от яйцекладной пластинки последнего, от её заднего края, отходит третья пара створок, и все три пары створок объединяются в трубчатый яйцеклад. Первая пара располагается над второй парой створок и нередко сливается в непарный желобчатый орган, а третья пара иногда преобразуется в кроющее устройство, не принимающее участия в работе самого яйцеклада. Однако у прямокрылых третья пара створок входит в состав яйцеклада как деятельный компонент и вместе со створками второй пары соединяется скользящим швом со створками первой пары.
Приведённый в рабочее положение яйцеклад сверчков упирается своим острым концом в субстрат и затем за счёт попеременных движений створок, легко скользящих друг относительно друга, внедряется в его глубину. Эти движения обеспечиваются мышцами яйцекладных пластинок, причём первая из них имеет треугольную форму и гибко связана своей вершиной с соответствующей створкой (рис. 53).

Рисунок 53. Яйцеклад сверчка (по Шванвичу, 1949):
фр — фрагма девятого сегмента брюшка; я. пл I, я. пл II — первые и вторые яйцекладные пластинки; я. ств I, я. ств III — первая и третья створки яйцеклада
Качаясь на сочленении с фрагмой тергита девятого сегмента, она упирается верхним углом во вторую яйцекладную пластинку и через соответствующее сочленение приводит её в движение. При этом начинает двигаться и связанная с ней третья яйцекладная створка. Двигаясь во взаимно противоположных направлениях, обе пары створок проталкивают выходящее из яйцеводов яйцо по направлению к вершине яйцеклада.
Яйцеклад саранчовых, сохраняя общий план строения, становится орудием копания земли. Его основу составляют короткие, изогнутые в виде крючьев первая и третья пары створок, тогда как вторая пара створок невелика и помещается между ними. Сдвигая и раздвигая створки, самка саранчи прокладывает глубокие ходы даже в твёрдом грунте; при этом её брюшко растягивается, увеличиваясь в длину в 2–3 раза (до 10 см). Таким образом, откладываемые ею яйца оказываются погребёнными в почве, а выделяющийся секрет образует вокруг них плотную кубышку с затвердевающими, как цемент, стенками.
Сходным образом устроены яйцеклады многих других насекомых: тараканов, клопов, цикад и перепончатокрылых. У рогохвостов и пилильщиков, откладывающих яйца в столь твёрдые субстраты, как древесина, яйцекладные створки снабжены рёбрышками и зубцами, что позволяет им работать в качестве пил и напильников. Своеобразны яйцеклады наездников, во много раз превосходящие длину тела, и утратившее свою функцию жало ос, пчёл, шмелей. Так, у медоносной пчелы яйцекладные пластинки, именуемые треугольной и продолговатой, связаны посредством сочленений друг с другом и с тергитом девятого сегмента (рис. 54).

Рисунок 54. Жало пчелы (по Weber, 1966):
я. пл I, я. пл II — первая и вторая яйцекладные пластинки; я. ств I, я. ств II, я. ств III — первая, вторая и третья яйцекладные створки; трг IX — терпит девятого сегмента брюшка
Совершая качания, они обеспечивают скольжение створок друг относительно друга. Створки третьей пары выполняют роль ножен и непосредственно перед нанесением укола отгибаются вверх, обнажая жало. Створки второй пары расширены у основания в барабан или луковицу, внутри которой, как поршень насоса, перемещаются отростки створок первой пары. Связанные с резервуаром ядовитой железы, эти структуры содействуют нагнетанию яда в место укола, а мелкие зазубринки створок обеспечивают углубление и фиксацию жала в коже жертвы. Вместе с тем эти зазубринки не позволяют пчеле вытащить жало из кожи; будучи спугнутой, она расстаётся со своим оружием и гибнет. Вырванное при этом жало сохраняет связь с оторванными вместе с ним резервуаром ядовитой железы и ганглием брюшной нервной цепочки. Именно поэтому жало, даже оторванное от тела, продолжает углубляться в ранку и впрыскивать новые порции яда. В отличие от пчёл осы не расстаются со своим жалом, лишённым зазубринок. Особенно опасны для человека крупные осы — шершни, сильный яд которых может привести к тяжёлому отравлению. Таким образом, яйцеклады, являясь производными конечностей брюшных сегментов, исполняют функции, часто не связанные непосредственно с откладкой яиц.
Строение копулятивных органов самцов насекомых выводится из исходного состояния щетинохвосток. Расположенный на фаллобазе эдеагус приобретает у высших насекомых разные формы, что часто используется в систематике для дискриминации видов. Причленённые с двух сторон к фаллобазе лопасти парамер (рис. 55), по-видимому, гомологичны третьим яйцекладным створкам. Эти структуры образуют фаллическую группу придатков, окружённую компонентами перифаллической группы. Составленные производными девятого брюшного сегмента — широкими лопастями вальв — они соответствуют грифелькам коксоподитов.

Рисунок 55. Схема строения копулятивных придатков самца (по Шванвичу, 1949):
цер — церки; ан — анальное отверстие; пар — парамеры; эд — эдеагус; фб — фаллобаза; пол — половое отверстие
Приведённая схема строения подвергается значительным модификациям, причём особенно резкие изменения испытывают половые сегменты самцов двукрылых. Нередко они становятся асимметричными или поворачиваются вокруг продольной оси тела на 180–360 °, так что стерниты могут оказаться сверху, а тергиты — снизу или занять не свойственные им боковые положения.

