10.8. Insects of the marine, intertidal, and littoral zones
The estuarine and subtropical and tropical mangrove zones are transitions between fresh and marine waters. Here, the extremes of the truly marine environment, such as wave and tidal actions, and some osmotic effects, are ameliorated. Mangroves and “saltmarsh” communities (such as Spartina, Sarcocornia, Halosarcia, and Sporobolus) support a complex phytophagous insect fauna on the emergent vegetation. In intertidal substrates and tidal pools, biting flies (mosquitoes and biting midges) are abundant and may be diverse. At the littoral margin, species of any of four families of hemipterans stride on the surface, some venturing onto the open water. A few other insects, including some Bledius staphylinid beetles, cixid fulgoroid bugs, and root-feeding Pemphigus aphids, occupy the zone of prolonged inundation by salt water. This fauna is restricted compared with freshwater and terrestrial ecosystems.
Splash-zone pools on rocky shores have salinities that vary because of rainwater dilution and solar concentration. They can be occupied by many species of corixid bugs and several larval mosquitoes and crane flies. Flies and beetles are diverse on sandy and muddy marine shores, with some larvae and adults feeding along the strandline, often aggregated on and under stranded seaweeds.
Within the intertidal zone, which lies between high and low neap-tide marks, the period of tidal inundation varies with the location within the zone. The insect fauna of the upper level is indistinguishable from the strandline fauna. At the lower end of the zone, in conditions that are essentially marine, crane flies, chironomid midges, and species of several families of beetles occur. The female of a remarkable Australasian marine trichopteran (Chathamiidae: Philanisus plebeius) lays its eggs in a starfish coelom. The early-instar caddisflies feed on starfish tissues, but later free-living instars construct cases of algal fragments.
Three lineages of chironomid midges are amongst the few insects that have diversified in the marine zone. Telmatogeton (Fig. 6.11) is common in mats of green algae, such as Ulva, and occurs worldwide, including many isolated oceanic islands. In Hawai’i the genus has re-invaded freshwater. The ecologically convergent Clunio also is found worldwide. In some species, adult emergence from marine rock pools is synchronized by the lunar cycle to coincide with the lowest tides. A third lineage, Pontomyia, ranges from intertidal to oceanic, with larvae found at depths of up to 30 m on coral reefs.
The only insects on open oceans are pelagic water-striders (Halobates), which have been sighted hundreds of kilometers from shore in the Pacific Ocean. The distribution of these insects coincides with mid-oceanic accumulations of flotsam, where food of terrestrial origin supplements a diet of marine chironomid midges.
Physiology is unlikely to be a factor restraining diversification in the marine environment because so many different taxa are able to live in inland saline waters and in various marine zones. When living in highly saline waters, submerged insects can alter their osmoregulation to reduce chloride uptake and increase the concentration of their excretion through Malpighian tubules and rectal glands. In the pelagic water-striders, which live on the surface film, contact with saline waters must be limited.
As physiological adaptation appears to be a surmountable problem, explanations for the failure of insects to diversify in the sea must be sought elsewhere. The most likely explanation is that the insects originated well after other invertebrates, such as the Crustacea and Mollusca, had already dominated the sea. The advantages to terrestrial (including freshwater) insects of internal fertilization and flight are superfluous in the marine environment, where gametes can be shed directly into the sea and the tide and oceanic currents aid dispersal. Notably, of the few successful marine insects, many have modified wings or have lost them altogether.
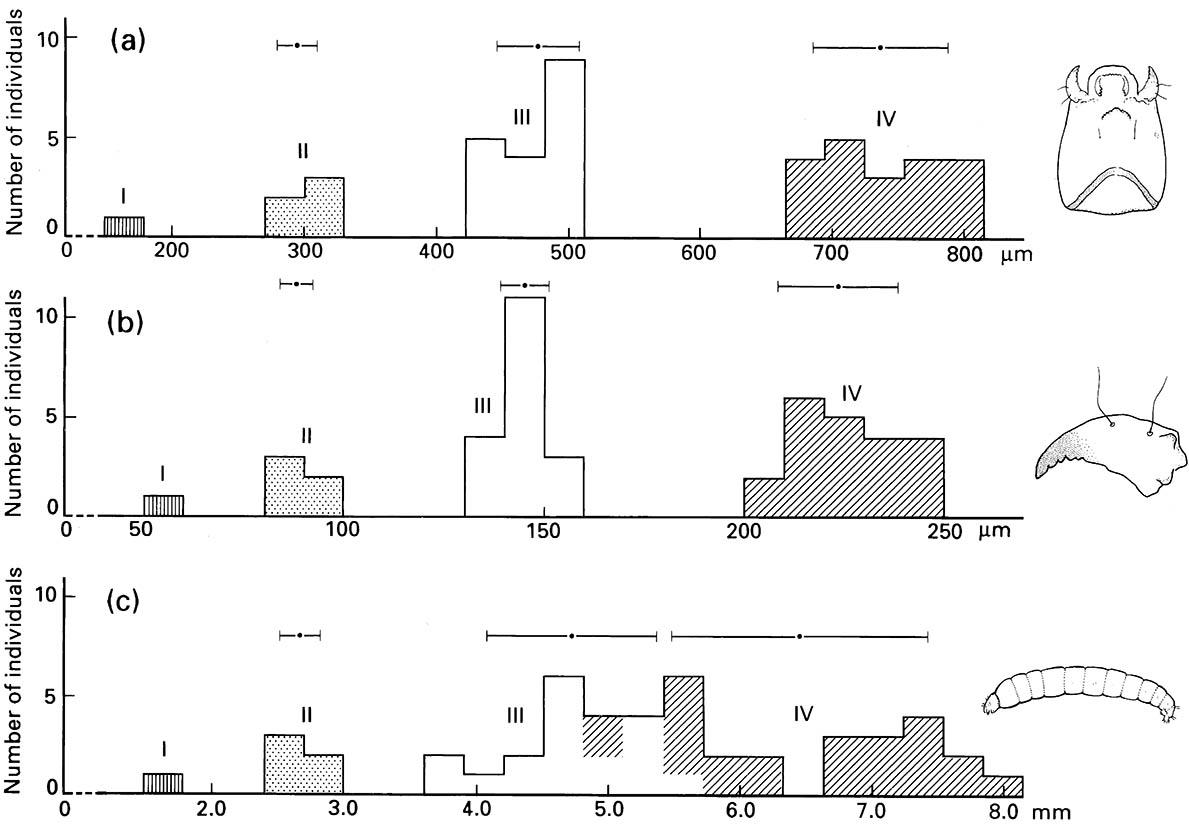
(a) head capsule length; (b) mandible length; and (c) body length between the four larval instars (I–IV). The dots and horizontal lines above each histogram represent the means and standard deviations of measurements for each instar. Note that the lengths of the sclerotized head and mandible fall into discrete size classes representing each instar, whereas body length is an unreliable indicator of instar number, especially for separating the third- and fourth-instar larvae.

