10.3.3. Oxygen uptake with a closed tracheal system
Simple cutaneous gaseous exchange in a closed tracheal system suffices for only the smallest aquatic insects, such as early-instar caddisflies (Trichoptera). For larger insects, although cutaneous exchange can account for a substantial part of oxygen uptake, other mechanisms are needed.
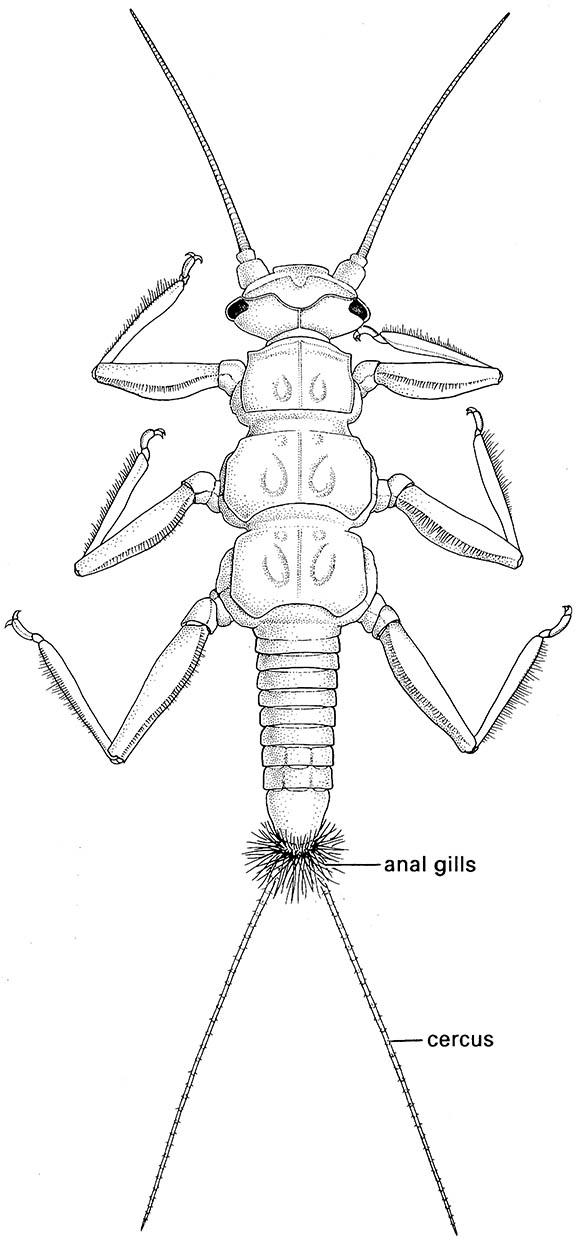
A prevalent means of increasing surface area for gaseous exchange is by gills — tracheated cuticular lamellar extensions from the body. These are usually abdominal (ventral, lateral, or dorsal) or caudal, but may be located on the mentum, maxillae, neck, at the base of the legs, around the anus in some Plecoptera (Fig. 10.1), or even within the rectum, as in dragonfly nymphs. Tracheal gills are found in the immature stages of Odonata, Plecoptera, Trichoptera, aquatic Megaloptera and Neuroptera, some aquatic Coleoptera, a few Diptera and pyralid lepidopterans, and probably reach their greatest morphological diversity in the Ephemeroptera.
In interpreting these structures as gills, it is important to demonstrate that they do function in oxygen uptake. In experiments with nymphs of Lestes (Odonata: Lestidae), the huge caudal gill-like lamellae of some individuals were removed by being broken at the site of natural autotomy. Both gilled and ungilled individuals were subjected to low-oxygen environments in closed-bottle respirometry, and survivorship was assessed. The three caudal lamellae of this odonate met all criteria for gills, namely:
- large surface area;
- moist and vascular;
- able to be ventilated;
- responsible normally for 20–30% of oxygen uptake.
However, as temperature rose and dissolved oxygen fell, the gills accounted for increased oxygen uptake, until the maximum uptake reached 70%. At this high level, the proportion equaled the proportion of gill surface to total body surface area. At low temperatures (<12°C) and with dissolved oxygen at the environmental maximum of 9 ppm, the gills of the lestid accounted for very little oxygen uptake; cuticular uptake was presumed to be dominant. When Siphlonurus mayfly nymphs were tested similarly, at 12—13°C the gills accounted for 67% of oxygen uptake, which was proportional to their fraction of the total surface area of the body.
Dissolved oxygen can be extracted using respiratory pigments. These pigments are almost universal in vertebrates but also are found in some invertebrates and even in plants and protists. Amongst the aquatic insects, some larval chironomids (bloodworms) and a few notonectid bugs possess hemoglobins. These molecules are homologous (same derivation) to the hemoglobin of vertebrates such as ourselves. The hemoglobins of vertebrates have a low affinity for oxygen; i.e. oxygen is obtained from a high-oxygen aerial environment and unloaded in muscles in an acid (carbonic acid from dissolved carbon dioxide) environment — the Bohr effect. Where environmental oxygen concentrations are consistently low, as in the virtually anoxic and often acidic sediments of lakes, the Bohr effect would be counterproductive. In contrast to vertebrates, chironomid hemoglobins have a high affinity for oxygen. Chironomid midge larvae can saturate their hemoglobins through undulating their bodies within their silken tubes or substrate burrows to permit the minimally oxygenated water to flow over the cuticle. Oxygen is unloaded when the undulations stop, or when recovery from anaerobic respiration is needed. The respiratory pigments allow a much more rapid oxygen release than is available by diffusion alone.