7.1.1. Systematic methods
The various methods that attempt to recover the pat- tern produced by evolutionary history rely on observations on living and fossil organisms. As a simplification, three differing methods can be identified: phenetics, cladistics, and evolutionary systematics.
The phenetic method (phenetics) relies on estimates of overall similarity, usually derived from morphology, but sometimes from behavior and other traits, and increasingly from molecular evidence. Many of those who have applied phenetics have claimed that evolution is unknowable and the best that we can hope for are patterns of resemblance; however, other scientists believe that the phenetic pattern revealed is as good an estimate of evolutionary history as can be obtained. Alternative methods to phenetics are based on the premise that the pattern produced by evolutionary processes can be estimated, and, furthermore, ought to be reflected in the classification. Overall similarity, the criterion of phenetics, may not recover this pattern of evolution and phenetic classifications are therefore artificial.
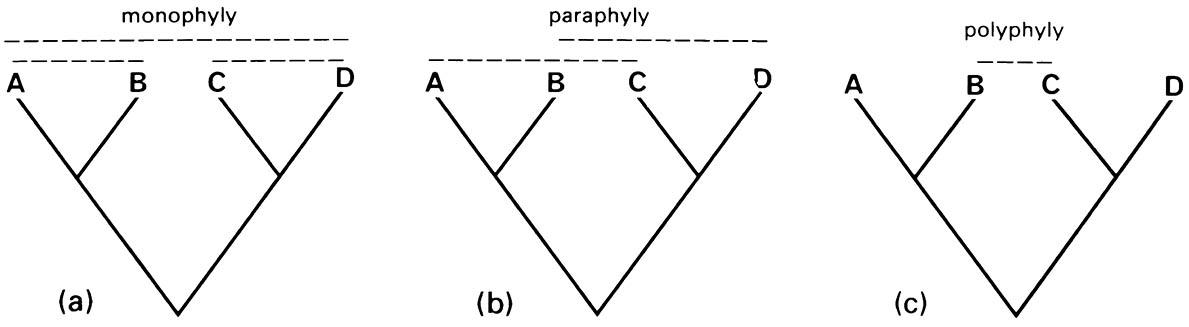
The cladistic method (cladistics) seeks patterns of special similarity based only on shared, evolutionarily novel features (synapomorphies). Synapomorphies are contrasted with shared ancestral features (plesiomorphies or symplesiomorphies), which do not indicate closeness of relationship. Furthermore, features that are unique to a particular group (autapomorphies) but unknown outside the group do not indicate inter-group relationships, although they are very useful for diagnosing the group. Construction of a cladogram (Fig. 7.1), a treelike diagram portraying the phylogenetic branching pattern, is fundamental to cladistics. From this tree, monophyletic groups, or clades, their relationships to each other, and a classification, can be inferred directly. Sister groups are taxa that are each other’s closest relatives. A monophyletic group contains a hypothetical ancestor and all of its descendants.
Further groupings can be identified from Fig. 7.1: paraphyletic groups lack one clade from amongst the descendants of a common ancestor, and often are created by the recognition (and removal) of a derived subgroup; polyphyletic groups fail to include two or more clades from amongst the descendants of a common ancestor (e.g. A and D in Fig. 7.1c). Thus, when we recognize the monophyletic Pterygota (winged or secondarily apterous insects), a grouping of the remainder of the Insecta, the non-monophyletic “apterygotes”, is rendered paraphyletic. If we were to recognize a group of flying insects with wings restricted to the mesothorax (dipterans, male coccoids, and a few ephemeropterans), this would be a polyphyletic grouping. Paraphyletic groups should be avoided if possible because their only defining features are ancestral ones shared with other indirect relatives. Thus, the absence of wings in the paraphyletic apterygotes is an ancestral feature shared by many other invertebrates. The mixed ancestry of polyphyletic groups means that they are biologically uninformative and such artificial taxa should never be included in any classification.
Evolutionary systematics also uses estimates of derived similarity but, in contrast to cladistics, estimates of the amount of evolutionary change are included with the branching pattern in order to produce a classification. Thus, an evolutionary approach emphasizes distinctness, granting higher taxonomic status to taxa separated by “gaps”. These gaps may be created by accelerated morphological innovation in a lineage, and/or by extinction of intermediate, linking forms. Thus, ants once were given superfamily rank (the Formicoidea) within the Hymenoptera because ants are highly specialized with many unique features that make them look very different from their nearest relatives. However, phylogenetic studies show ants belong in the superfamily Vespoidea, and are given the rank of family, the Formicidae (Fig. 12.2).
Current classifications of insects mix all three practices, with most orders being based on groups (taxa) with distinctive morphology. It does not follow that these groups are monophyletic, for instance Blattodea, Psocoptera, and Mecoptera almost certainly are each paraphyletic (see below). However, it is unlikely that any higher-level groups are polyphyletic. In many cases, the present groupings coincide with the earliest colloquial observations on insects, for example the term “beetles” for Coleoptera. However, in other cases, such old colloquial names cover disparate modern groupings, as with the old term “flies”, now seen to encompass unrelated orders from mayflies (Ephemeroptera) to true flies (Diptera). Refinements continue as classification is found to be out of step with our developing understanding of the phylogeny. Thus, current classifications increasingly combine traditional views with recent ideas on phylogeny.

The superfamily Apoidea includes the Sphecidae sensu stricto, the Crabronidae (formerly part of a broader Sphecidae), the Ampulicidae (not shown), and all bees, here treated as one family, the Apidae, with several subfamilies (e.g. Apinae, Colletinae, Halictinae; not all solitary groups are shown) of uncertain relationships. Traditionally, bees have been classified in several families, a ranking that is unjustified phylogenetically. Probable relationships within non-social aculeate wasps (e.g. Ampulicidae, Pompilidae, and Rhopalosomatidae) and bees are not depicted. (Adapted from several sources including Gauld & Bolton 1988; Alexander 1992; Brothers 1999; B.N. Danforth, pers. comm.)

